メンデルの遺伝について
世代から世代への遺伝子の伝達
| このページの内容 |
遺伝:形質はどのように遺伝するのか?
- 1822 年にメンデル Johann Gregor Mendel はハインツェンドルフ(現,チェコスロバキア領ハインツァイス)で生まれた。彼は学生として将来を有望視されていたが,貧困のため大学に進学することができなかった。
- 1843 年 21 歳の時,博物学の興味を捨てきれず,ブリュン(現,ブルノ Brno) のアウグスティノ修道院に入った。
- 1851 年の秋に,すなわち修道士としての修行が終わった後,メンデルはウィーン大学に留学した。1 年目に物理学,数学,化学ならびに自然科学の科目を受けた。植物学の科目で,メンデルはすべての生物は細胞からできており,細胞がすべての生物の基本単位であるとする新しい概念に出合った。
この細胞説は,いくつかの新しい問題と多くの矛盾する問題を示した。問題の 1 つは,両親が子供の形質に均等に影響を与えているのかということであった。- ほとんどの動植物で雌性配偶子は雄性配偶子よりも大きいので,これは当然の疑問であり,議論の多い問題であった。
- これに関連する問題として,子供の形質は両親の形質が混じり合って受け継がれるのかどうかということである。
もしそうならば,女性の貢献度は 40% なのか,50% なのか,60% なのか,あるいはそれ以上なのか謎だらけであった。
- 1854 年メンデルは Brno に戻り物理学を教えながら,これらの疑問を解くための実験の準備を始めた。
| 閑話休題~メンデルと試験不安症 |
|---|
|
| もし,あなたが「家畜育種学」の講義や試験を受けている時にストレスを感じたら,メンデルにとってはもっと酷かったのだと思いなおして,気分を楽にして欲しい。 |
メンデルの実験は多くの疑問を解決した
メンデルが遺伝の仕組みを解明できたのは,綿密に計画された実験の結果である。研究目的を明確にしてから,メンデルは用いる生物を選択している。このことについては彼の画期的な論文のはじめの方で述べられている。
- ” 実験の価値や正当性は,用いられた方法が適切であるかどうかで決まる。本研究でも,どの植物を実験に選択するか,またどうのように行うかということが最も重要である。成功の可能性をすべて無駄にしないために,この種の実験では慎重に植物の種類が選択されるべきである。”
そこで,彼は実験用生物に期待される特性のリストを作った。
- まず,研究すべき 異なる形質を多く もっていること
- 植物は自家受精し,偶発的な受粉が起こりにくい花の構造をもっていること
- 自家受粉した子孫は繁殖力があり,次世代の交配が可能であること
彼はとくに エンドウ の仲間に興味を持った。その理由は,偶発的な受粉が起こりにくい花の構造をしていたからである。 彼は,エンドウのいろいろな形質をもった 34 品種を種子業者から手に入れた。この植物は,
{kind=link}
- 比較的短期間に生長し,
- 地蒔きでも,温室のポット栽培でも発育でき,
- さらに自家受粉も人工授粉も可能であった。
| 表 1. メンデルの研究に用いられた形質 | ||
|---|---|---|
| 形質 | 優性 | 劣性 |
| 種子 | ||
| 形状 | 丸種 | しわ種 |
| 色 | 黄色 | 緑色 |
| 種皮色 | 灰色 | 白色 |
| 鞘 | ||
| 形状 | 膨らみ | くびれ |
| 色 | 緑色 | 黄色 |
| 花 | ||
| 配置 | 葉腋 ( ようえき ) | 先端 |
| 茎 | ||
| 長さ | 長い | 短い |
メンデルは,エンドウのもっている同一の形質が自家受粉によってすべての子孫に代々伝わるのかを 2 年にわたり 34 品種のすべてについて確認のための実験をしている。この実験から,彼は 22 品種を選抜して,本試験として 8 年の間毎年植えている。[ メンデルが実験をした修道院の庭 ]
{kind=link}
そして,種子,鞘,花ならびに茎の構造に影響する 7 種類の形質を見出している( 表 1 )。
各形質は 2 つの明瞭な形態によって区別できた。たとえば,茎が高いものと低いもの,種子の形状がしわがあるもの ( しわ種 ) と表面が平滑なもの ( 丸種 ) などである。
例数が少ないために起こる過誤をさけるために,彼は大規模の実験を計画した。さらに 8 年を要して,メンデルは 28,000 を超えるエンドウを実験に用いている。
彼は数世代にわたりそれぞれの形質を別々に追跡した。彼は,一対の形質毎に検討し始め,さらにその結果を確証するために追試している。
物理学や数学の教育を受けていたので,メンデルはデータを確率と統計の基礎に基づいた分析をしている。
実験に対して綿密に検討された研究手法と先入観をもたなかったことがメンデルの成功の秘訣であった。
エンドウの交配:単一形質の関するメンデルの実験

メンデルが遺伝についてどのように考えていったのかを知るために,まず彼が得た実験結果のいくつかをみてみよう。それから,メンデルが結論に達する論理的思考と,さらにその発想を確認するための実験について述べる。最初の実験では,種子形状の遺伝について検討している。この場合 1 つの形質のみを問題にしているので,彼はこれを 一因子交雑 monohybrid cross と呼んだ。
| 一因子交雑試験の要約版はこちら |
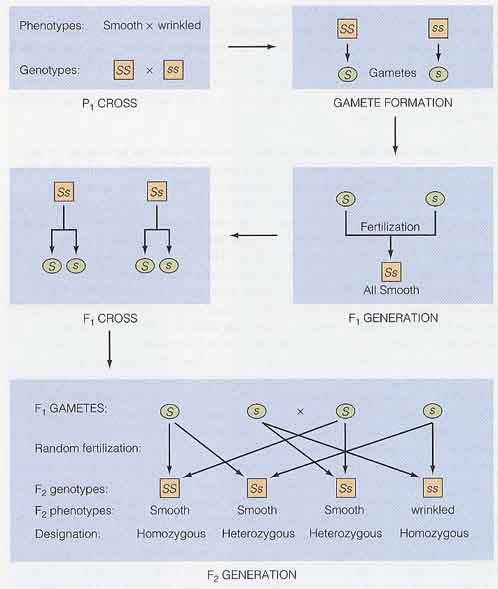
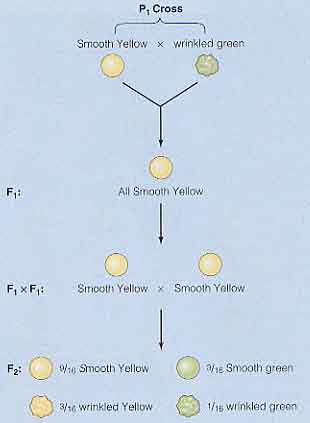
彼は丸種をもつエンドウを選び,しわ種をもつものと交雑した。交雑を確実にするために,人工授粉を行っている。
この最初の実験で,メンデルは 15 品種に 60 回受粉をさせ,その結果生じた種子はすべて丸種であった。しわ種のものを授粉しても丸種のものを授粉しても結果は変わらなかった。
翌年,メンデルはこの交雑で得た丸種を植えた。エンドウが生長し自家受粉して,7,324 個の種子を回収した。これらのうち,5,474 個が丸種で,1,850 個がしわ種であった。
- P1 : 丸種×しわ種
- F1 : すべて丸種
- F2 : 5,474 個が丸種,1,850 個がしわ種
メンデルはこの実験で,親世代を P1 と呼び,子供を F1 ( first filial )世代と名付けた。F1 個体の自家受粉によって生産された第 2 世代を F2 ( second filial ) 世代と呼んだ。種子形状に対するメンデルの実験を右図にまとめた。
メンデルの最初の交雑試験の結果と結論
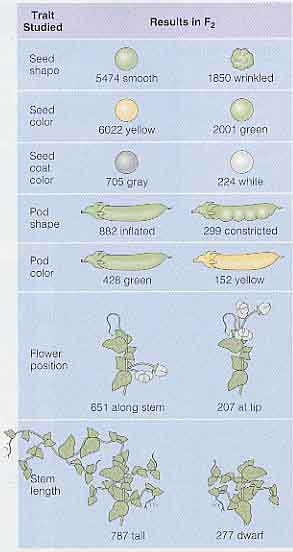 7 つの形質すべてに関する実験の結果は,メンデルが丸種としわ種について観察したものと同様であった( 右図 )。すべての交雑の結果は以下の通りであった。
7 つの形質すべてに関する実験の結果は,メンデルが丸種としわ種について観察したものと同様であった( 右図 )。すべての交雑の結果は以下の通りであった。
- F1 世代は親の形質のいずれかを示し,必ず ( 親と ) 同じ形質が出現した。
- すべての交雑で,どの植物の花粉と受粉したかは全く影響がなかった。結果は常に同一であった。
- F1 世代で観察されない形質は F2 世代の約 25% に発現した。
これらの結果は,メンデルの最初の発見であった。彼の実験は,形質が全く変化しないまま親から子へ伝わり,形質の発現が無くても,世代を超えて変化しないまま伝わることを示した。
このことから,メンデルは親の形質が子供に混じり合って伝わるのではないことを確信した。むしろ,形質は混じり合うことがない分離した単位で遺伝すると結論した。
メンデルは正逆交配を実験的に行った。したがって,ある実験で雄性親として用いた品種を,次の交雑試験では雌性親として用いた。すべての例で,雄性か雌性かに関わらず結果は同一であった。これらの実験から,各親は子供に対して遺伝的に等しく貢献していると結論した。
7 つの形質のそれぞれについて交雑した結果をもとに,メンデルは次のような結論を導き出した。
- 形質を決定する 遺伝子 ( メンデルは 因子 と呼んでいた ) は発現しないことがある。
たとえば,丸種の個体としわ種の個体を交配した場合,すべての F1 種子は丸種であった。これらの種子を育て,自家受粉させると次世代 ( F2 ) ではしわ種のものが含まれる。このことは,F1 種子がしわ種になる遺伝子 ( 存在するが,発現しない遺伝子 ) をもっていることを意味する。彼は F1 で発現せず,F2 世代で発現する形質を 劣性形質 recessive trait と呼び,F1 世代で発現する形質を 優性形質 dominant trait と呼んだ。 - 丸種の P1 の子孫と丸種の F1 世代を比較することによって,外見は同じにもかかわらず,P1 と F1 では遺伝的に異なるとメンデルは結論づけた。
P1 が自家受粉すると,次世代のすべての個体は丸種だけである。しかし,F1 を自家受粉すると F2 の個体は丸種としわ種の両方が出現する。メンデルは生物の外見とその遺伝的構成との間に違いがあること,そしてそれを明確にすることが重要であることを完全に理解していたのである。現在では,
- 我々が目にする形質をいう時には 表現型 phenotype
- 生物の遺伝的構成をいう時には 遺伝子型 genotype
という用語が使われている。上記の例では,丸種をもつ P1 と F1 では表現型は同一であるが,両者の遺伝子型は異なるということである。
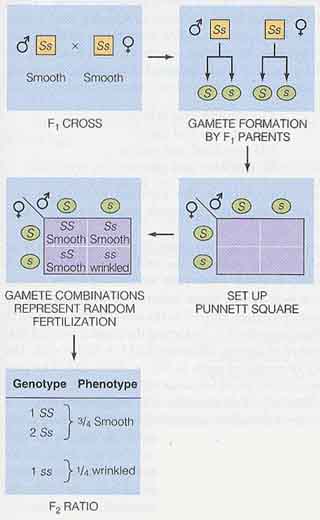
- これらの自家受粉の実験結果から,F1 個体は丸種形質としわ種形質の遺伝子をもつことが示される。
そこで,F1 個体には種子形状を支配する遺伝子がいくつあるのかという疑問が生じてくる。雄性と雌性の親が,子供の形質に均等に貢献することをメンデルはすでに論理的に示している。
最も簡単な説明をすると,各 F1 個体は 2 個の遺伝子をもち,うち 1 個は丸種形質を発現し,もう 1 個は発現しないままのしわ種形質の遺伝子である。この論理を拡張することによって,P1 のそれぞれの個体と F1 も種子形状に関する 2 個の遺伝子をもつ。遺伝子を記号で表わすために,大文字を優性形質の遺伝子を表わすのに,小文字は劣性形質を表わすのに用いられる ( S,丸種;s,しわ種 )。この省略表現を用いると,P1 と F1 の遺伝子型と表現型が このように 表わすことができる。
{kind=link}
1 つの形質はどのように遺伝するのか:分離の法則
遺伝子が対で存在すると,世代が進む時にそれらが倍加しないような仕組みがあるはずである ( 各親がある形質の遺伝子を 2 個もっていたとすると,なぜ子供では 4 個にならないのかということである )。遺伝子対のそれぞれが配偶子形成の時,お互いに分離するとメンデルは推論した。結果として各配偶子はある形質を制御する 2 個の遺伝子の 1 個を受け継ぐことになる。配偶子形成時の遺伝子対の分離は 分離の法則 the principle of segregation と呼ばれている。
遺伝子対の分離の様子を右図に示す。F1 世代の各構成員は同じ割合で 2 種類の配偶子をつくる ( S 配偶子と s 配偶子 )。受精時の配偶子の任意な組み合わせが,ピュネット・スクエア Punnett square ( R.C. Punnett によって考案された遺伝子交雑を分析するための方法 ) で示すような遺伝子型の組み合わせを生む。
F2 は,1SS : 2Ss : 1ss の遺伝子型比と,3 丸種 : 1 しわ種の表現型比をもつ。
メンデルの論理的な考え方によって,F2 世代の遺伝子型が予測できるようになった( 右図 )。
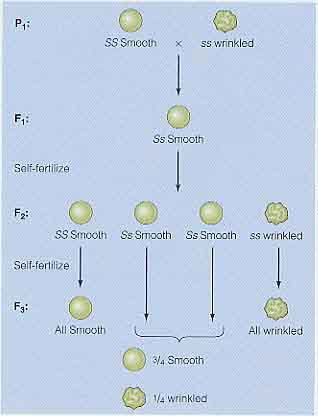
- F2 個体の 1/4 が丸種の遺伝子 ( SS ) だけをもっており,自家受粉させるとすべての個体は丸種をもつ。
- F2 個体の 1/2 は丸種としわ種の両方の遺伝子をもつはずである ( Ss )。そして,自家受粉させて F3 世代を生産すれば 3 : 1 の割合で丸種としわ種が生じる。
- 最後に残った F2 個体の 1/4 はしわ種の遺伝子のホモ接合体 ( ss ) で,自家受粉させてもすべてがしわ種の子孫が得られる。
実際,メンデルは F2 世代の多数の個体を受精させ, 5 世代まで連続してこの予測したが正しいかを検証している。
| メンデルは,体細胞分裂と減数分裂が発見させる前に,あるいはまた染色体が発見される前に実験を始めている。後述するように,形質がどのように遺伝するのかということに関するメンデルの結論は,事実,減数分裂における染色体の動態を説明しているのである。この点をみただけでも,メンデルの発見には目を見張るものがある。 |
今日我々は,”メンデルの因子” を 遺伝子 と呼び,それぞれ組み合わされる遺伝子を 対立遺伝子 allele と呼んでいる。既にみてきた例では,種子形状の遺伝子は 2 個の対立遺伝子,すなわち丸種遺伝子 S としわ種遺伝子 s をもつ。
全く同じ対立遺伝子をもつ個体はホモ接合体 ( SS と ss ) であり, 2 つの異なる対立遺伝子が存在する場合はヘテロ接合体の遺伝子型 ( Ss ) をもつ。SS ホモ接合体と Ss ヘテロ接合体は優性の丸種の表現型を示し,ss ホモ接合体は劣性のしわ種表現型を示す ( S は s に対して優性である )。
さらに多くの交雑が行われた:独立の法則
メンデルは一因子交雑から,より複雑な状況の遺伝について検討する必要があることに気がついていた。彼は論文の中で次のように述べている:
- ” これまで述べた実験では, 1 つの形質だけが異なる植物を用いた。ここで見出した法則が,異なる 2 つの形質を組み合わせても適応できるかどうかを調べることが次の実験の目的である。”
この目的のために,彼は種子の 「 形状 」 と 「 色 」 を研究対象とした。彼は 2 つの形質を含む交雑を 二因子交雑 dihybrid cross と呼んだ。
二形質を含む交雑
メンデルが実際に行った実験の結果についてみてみよう。これまで行った交雑実験で,メンデルは種子については,丸種がしわ種に対して優性であること,また黄色が緑色に対して優性であることを知っていた。ここでは,それぞれを丸種 ( S ),しわ種 ( s ),黄色 ( Y ),緑色 ( y ) で表わす。
結果と結論
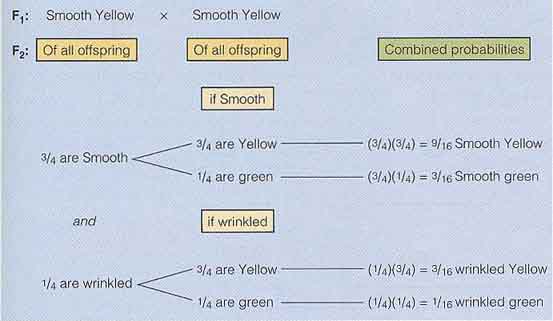
 メンデルは丸種で黄色の種子をもつ純粋種を選抜し,それらをしわ種で緑色の種子をもつ純粋種に交雑した( 右図 )。F1 個体では,種子はすべて丸種で黄色であった。このことから,丸種と黄色が優性形質であることが確認された。そこで,メンデルは F1 個体を自家受粉させ,F2 世代を生産した。これらの F2 個体は 4 種類の種子を生産し, 1 つの鞘の中に混在していた。15 の個体から,メンデルは合計 556 個の以下のような種子を得た。
メンデルは丸種で黄色の種子をもつ純粋種を選抜し,それらをしわ種で緑色の種子をもつ純粋種に交雑した( 右図 )。F1 個体では,種子はすべて丸種で黄色であった。このことから,丸種と黄色が優性形質であることが確認された。そこで,メンデルは F1 個体を自家受粉させ,F2 世代を生産した。これらの F2 個体は 4 種類の種子を生産し, 1 つの鞘の中に混在していた。15 の個体から,メンデルは合計 556 個の以下のような種子を得た。
- 丸 種,黄 色 315 個
- 丸 種,緑 色 108 個
- しわ種,黄 色 101 個
- しわ種,緑 色 32 個
F2 世代の表現型には,P 世代の表現型や全く新しい表現型 ( 丸種,緑色としわ種,黄色 ) が含まれていた。二因子交雑で 2 個の遺伝子がどのように遺伝するのかを決定するために,メンデルはまず F2 世代の結果をそれぞれの形質に分けて分析した ( この場合, 2 形質以外の形質は考慮していない )( 下図 )。
- 種子の色は無視して形状だけをみた場合には,F2 世代では 3/4 が丸種で, 1/4 がしわ種になるはずである。
丸種を示した個体の合計は 315 + 108 = 423 で,しわ種の合計は101 + 32 = 133 であった。丸種のしわ種に対する割合は 3 : 1 に極めて近い。 - 同様に,種子色だけを考慮すると,F2 世代では 416 個の黄色 ( 315 + 101 ) と 140 個の緑色 ( 108 + 32 ) がみられた。
これらも 3 : 1 に非常に近い。
それぞれの形質を分けると 3 : 1 の割合になること ( 分離の法則と同じ ) を確認してから,メンデルは 2 つの形質の遺伝を同時に検討した。
二形質の遺伝:独立の法則
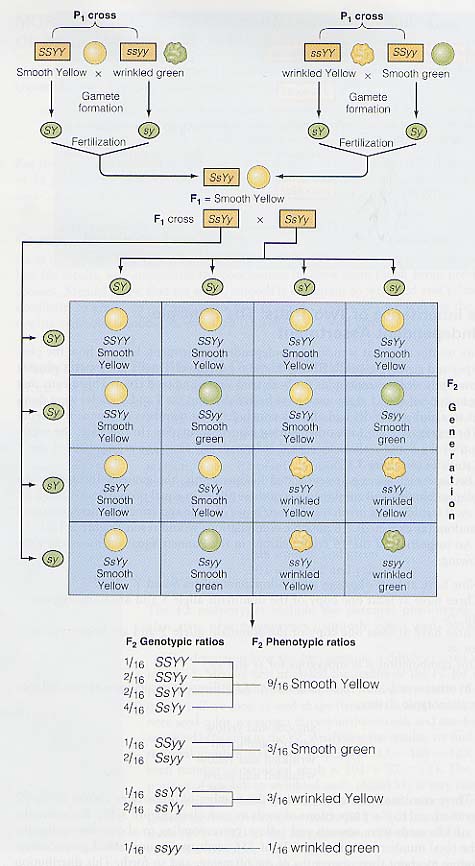
独立の原則について考える前に,F1 と F2 の表現型と遺伝子型がどのように生じるかをみてみよう。丸種をもつ F1 植物体は,種子形状も色も共にヘテロ接合体であった。このことは,F1 個体の遺伝子型が,SsYy で,遺伝子 S と Y はそれぞれ s と y に対し優性であることを示す。
メンデルはすでに遺伝子対のそれぞれが配偶子形成時に分離することを明らかにしていた。この場合,遺伝子 S と s の分離は遺伝子 Y と y の分離と独立に起こっていた。 [ 二因子実験のピュネット・スクエア はこちらを参照のこと ]
{kind=link}
遺伝子対のそれぞれが独立して分離するので,F1 個体によって形成される配偶子はこれらの対立遺伝子のすべての組み合わせを均等にもつ。すなわち,SY,Sy,sY,sy である。 4 種類の雄性ならびに雌性配偶子による受精が無作為に起こるとすると,16 通りの組み合わせが可能である ( 右図 )。
ピュネット・スクエアで 16 通りの組み合わせを検査すると,次のようなことが明らかになる。
- 優性遺伝子 S または Y を少なくとも 1 個もつのは 9 通りある。
- 優性遺伝子 S を少なくとも 1 個と yy のホモをもつのは 3 通りある。
- 優性遺伝子 Y を少なくとも 1 個と ss のホモをもつのは 3 通りある。
- ss と yy のホモの組み合わせは 1 つである。
言い換えると,16 通りの受精によって生じる遺伝子型が 4 種類の表現型にまとめることができる。
- 丸 種で黄 色
- 丸 種で緑 色
- しわ種で黄 色
- しわ種で緑 色
これらの組み合わせは F2 世代における表現型の数と,各表現型における種子の割合と一致する。 [ 詳細については 二因子実験のピュネット・スクエア を参照のこと ]たとえば,
- 556 個の種子のうち 315 個が丸種で黄色であった。これは全子孫数の 9/16 に相当する。
- また,556 個の種子のうち 108 個がしわ種で緑色であった。これは全体の 3/16 に相当する。
- 以下同様である。
F2 世代における分布は表現型比 9 : 3 : 3 : 1 となる。この交雑の結果は, ( メンデルがしたように ) 配偶子形成時に遺伝子対のそれぞれが独立して分離すると仮定するとよく説明できる。
このことから,すべての対立遺伝子の組み合わせを含んだ配偶子が形成されると結論できる。これがメンデルによって示された基本的な遺伝の原理で,独立の法則 と呼ばれている。
数千を超えるエンドウを用いた 10 年にも及ぶ研究の後,メンデルはその成果を 1865 年に彼が所属する Natural Science Society の 2 月と 3 月の学会で発表した。
発表の内容は翌年,同学会の会報に掲載された。彼の研究はいくつかの参考目録に紹介され,また論文のコピーが配布されたが,メンデルの発見の重要性の真価は認められなかった。
自然淘汰における遺伝の役割について触れたチャールズ・ダーウィンでさえ,メンデルの研究の重要性に気がつかなかった。メンデルの論文が発表された学会報のコピーはダーウィンも所有しており,いくつかの論文の余白には多くの書き込みが残されていたが,メンデルの論文のどこにも鉛筆の後が無かった。
そして1900年に,遺伝の機構についてそれぞれ独自に研究していた 3 人の科学者が,メンデルの発見を確証し,彼の論文が日の目を見たのであった。その結果,今日遺伝学 genetics と呼ばれている分野が大いに注目されたのであった。不幸にも,メンデルは 1884 年に死去しており,彼自身が遺伝学の基礎を打ち立てたことを知るよしもなかった。
減数分裂はメンデルの観察結果を説明する:遺伝子は染色体上にある
メンデルがエンドウで実験をしていた時,体細胞分裂ならびに減数分裂における染色体の動態については全く知られていなかった。
| 表 2. 遺伝子,染色体,ならびに減数分裂 | |
|---|---|
| 遺伝子 | 染色体 |
| 対立遺伝子として対を構成する | 相同遺伝子として対を構成する |
| 減数分裂時にお互いに分離した 遺伝子対からなる |
減数分裂時にお互いに分離した 相同染色体対からなる |
| 減数分裂時に独立に組み合わさ れた遺伝子対からなる |
減数分裂時に独立に組み合わさ れた染色体対からなる |
1900 年までに,それらの詳細が良く知られるようになった。メンデル遺伝の法則がすべての生物に当てはまることが確認されると,遺伝子と染色体が非常に共通点が多いことが明らかになってきた( 表 2 )。
- 染色体と遺伝子は 2 つ 1 組で存在する。
- 減数分裂の際には,染色体対のそれぞれがお互いに分離して,遺伝子対のそれぞれは配偶子形成時に分離する。
- そして最終的に,受精が起こり配偶子が融合して,2 倍体の染色体に戻る。
- 接合体に含まれる 2 セットの遺伝子が次世代の遺伝子型を形成する。
1903年に,W. サットン Walter Sutton と T. ボベリ Theodore Boveri もそれぞれ同様のことに気づき,遺伝子は染色体上あるとする考え方をそれぞれ独自に提唱した。この染色体説はその後多くの実験で確証され,近代遺伝学の基礎の一つとなった。各遺伝子は染色体上の特別な部位 ( 遺伝子座 locus ) を占め,各染色体は多くの遺伝子を運ぶ。ヒトでは,約 32,000 の遺伝子が 24 本の異なる染色体に乗っている。
| 減数分裂時に,遺伝子と染色体がどのように動くのかを示したページは こちらを参照のこと。 |
多くの遺伝子は 2 個以上の対立遺伝子をもつ
これまで我々は 2 個の対立遺伝子をもつ遺伝子だけを議論してきた。対立遺伝子は遺伝子の異なる型を表わすので,遺伝子が 2 個の対立遺伝子しかもたないという理由は無い。事実,多くの遺伝子が 2 個以上の対立遺伝子をもつ。どの個体も 1 遺伝子当たり 2 個の対立遺伝子をもつが,集団では 1 遺伝子に対して多数の対立遺伝子が存在する。
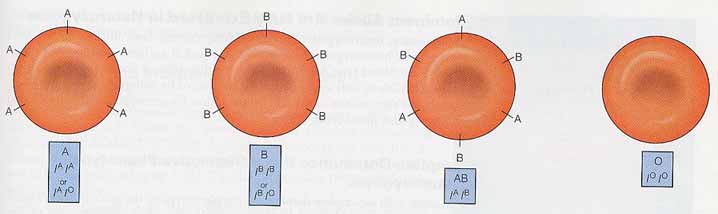
ヒトでは,ABO 式血液型を決定する遺伝子が 複対立遺伝子 multiple alleles をもつ遺伝子の良い例である。ABO 血液型はヒト赤血球の表面にある分子 ( 糖タンパク質 ) によって決定される。これらの分子が体の免疫系によって識別される細胞の認識票となる。
ABO 式血液型には, 3 個の対立遺伝子 IA,IB および IO をもつ 1 つの遺伝子( I )がある。対立遺伝子 A と B により,赤血球表面の分子の形状は僅かに異なって形成される。
A 遺伝子のホモ接合体 ( AA ) は細胞表面に A 抗原をもち,血液型が A 型となる。 同様に, B 遺伝子のホモ接合体 ( BB ) は細胞表面に B 抗原をもち,血液型が B 型となる。 3 つ目の対立遺伝子 O は抗原をつくらない。そしてこの遺伝子をホモでもつ人は細胞上に A 抗原も,B 抗原ももたない。対立遺伝子 O は対立遺伝子 A と B に対して劣性である。3 個の対立遺伝子があるため,遺伝子型は 6 組存在する( 下図 )。
メンデルの遺伝法則の拡張
メンデルの研究が広く知られるようになってから,F1 個体の表現型が両親のそれと似ていない例が見出されている。 ある例では,F1 個体が両親の表現型の中間型を示すものや,両親の両方の形質が現れる表現型を示すのもがみられた。そのため,メンデル遺伝によって説明可能なのか,あるいは分離や独立の法則によらない全く違う機構があるのかという議論が起こった。結局のところ,いくつか異なる生物を用いた実験でこれらの例がメンデル遺伝の例外ではないことが示された。この項ではこれらの変動の例と,表現型はメンデルの優性の法則に従わないが,遺伝子型の予測比はメンデルの法則に従うことを示す。
共優性対立遺伝子はヘテロ接合体でも完全に発現する
共優性の場合には,ヘテロ接合体でも両方の対立遺伝子が完全な発現を示す。ABO 式血液型で,AB ヘテロ接合体は赤血球膜上に A ならびに B 抗原の両方をもち,血液型は AB となる。この場合,どちらの対立遺伝子ももう一方に対して優性ではない。各対立遺伝子が完全に発現するので,共優性 codominance という( 上図参照 )。その結果,ABO 血液型の 3 つの対立遺伝子から遺伝子型の組み合わせ 6 通りでき,4 つの表現型が生じる。
不完全優性はヘテロ接合体で独特の表現型を示す

不完全優性が起こる場合には,ヘテロ接合体の表現型はホモ接合体の両親の表現型の中間型を示す。たとえば,スナップドラゴン( 右図 )では,赤い花を持つ純系の品種を,白い花を持つ品種と交配した場合には,F1 のすべてがピンクの花を持つ。F1 の表現型は両親のどちらとも異なり,その表現型の中間型である。F1 個体を自家受粉させた場合には,赤色,ピンク,白色の花を持つ F2 が 1 : 2 : 1 の割合で生産される。
この交配の結果はメンデルの法則で説明できる。
各遺伝子型はそれぞれ独特の表現型をもち,
表現型比 ( 1 赤色 : 2 ピンク : 1 白色 ) は
遺伝子型比 ( 1RR : 2Rr : 1rr ) と同一である。
赤色の花をつくるには,対立遺伝子 R が 2 個必要であり,1 個の場合にはピンクになってしまう。この場合,対立遺伝子 R は r に対して 不完全優性 であるという。対立遺伝子 r は色素をつくらないので,R 遺伝子がないと白色の花になるのである。
| 最初に戻る |
| メニューのページへ戻る |
March 11, 2020