組み換え型 DNA と遺伝子クローニング
![]()
25 April 2014 版を翻訳一部改変
| このページの内容 |
組み換え(型) DNA (recombinant DNA) は人工的に作られた DNA である。 2 つ以上の供給源から DNA を取り出し, 1 つの組み換え分子に結合することで形成された DNA 分子のことである。
組み換え recombinant DNA ( rDNA ) の作出 : 概要図
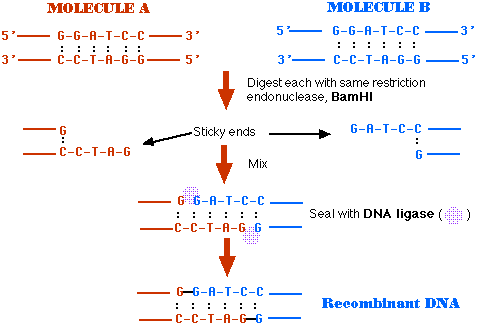
- 2 つの供給源から取り出した DNA を同じ 制限酵素 restriction endonuclease で処理する ( この例では BamHI として示してある ) 。
- BamHI は両方の DNA 分子の特異的な塩基配列を認識して,これを [G / GATCC] という風に切断する。
5′ GGATCC 3′
3′ CCTAGG 5′ - 切断されたDNA末端には 1 本鎖 DNA の突出した部分(突出末端)がある。
- これらは相補的な突出末端を含むDNA 分子と塩基対合するので,これらは 付着末端 sticky end と呼ばれる。
- この例では,両方の DNA 試料が相補的な付着末端をもつので,混合された時にお互いに塩基対合する。
- DNA リガーゼがこれら 2 つを共有結合させ, 1 つの 組み換え DNA 分子とする。
利用しやすいように, 組み換え分子は多数複製して,塩基配列を決定 などの分析の試料とする。同一組み換え分子のコピーを多数作製することを クローニング cloning と呼ばれ,通常 ポリメラーゼ連鎖反応 polymerase chain reaction ( PCR ) という技術が用いられる。しかし,ここでは生体内でどのようにクローニングが起こっているかをみてみよう。
以下の細胞で生体内でのクローニングが可能である。
すべての場合において, 複製され,発現され得る形態で組み換え DNA が細胞に取り込まれなければならない。これを実現するために,ベクター vector が使われ,DNA が導入されている。多くのウイルスが細菌や哺乳動物細胞に対するベクターとして使われている。ここでは,大腸菌を宿主として,プラスミド plasmid をベクターとしたクローニングの例を示す。
プラスミド plasmid
プラスミドは,細菌で見つかった染色体とは別個の DNA 分子である。プラスミドは:
- 非常に小さく ( 数千の塩基を含む ) ,
- 遺伝子を 1 または 2 – 3 個もち,
- 環状 で,
- 1 個の 複製起点 をもつ。
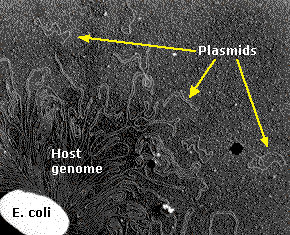
糸状の部分は,およそ4,300の遺伝子をコードしている460万以上の塩基対を含んでいる一本鎖DNA分子である。小さな輪状の部分はプラスミドを指す。
(ハーバード医科大学院のハンティントン・ポッターとデイビッド・ドレスラーの御好意による。)
プラスミドは細菌の染色体と同じ機構で複製される。プラスミドには,染色体と同じ割合でコピーされものと ( この場合は 1 個の細胞に 1 コピーのプラスミドが含まれる ) ,多量にコピーされるものがある ( この場合は 50 以上のコピーをもつことがある ) 。
多量のコピー数をもつプラスミドの遺伝子はたいてい高い割合で発現する。自然状態では,これらの遺伝子は 抗生物質 から細菌を護るタンパク質 ( 酵素など ) をコードしていることが多い。
プラスミドは細菌内に容易に入り込む。病院内などで抗生物質に抵抗性をもつ菌が急速に広がったりするのはこのためである。この性質を利用して,プラスミドは遺伝子導入に用いられる。
プラスミドの主な例
コールドスプリングハーバー研究所のデイビッド・ミクロスとグレッグ・フライアーの御好意による。彼らは,生物学に真剣に取り組んでいるすべての学生(高校生や大学生)が経験すべき実験基礎技術として,組み換えDNA技術へこれらのプラスミドを用いている。
pAMP
- 4539 個の塩基対
- 単一複製開始点
- 抗生物質 アンピシリン ampicillin ( 酸に安定で経口投与可能な広範囲ペニシリン ) 耐性の遺伝子 ( ampr )
- 以下の塩基の単独出現
5′ GGATCC 3′
3′ CCTAGG 5′この部分は上述のように 制限酵素 BamHI で切断できる。
- 以下の塩基の単独出現
5′ AAGCTT 3′
3′ TTCGAA 5′この部分は制限酵素 HindIII で切断できる。
BamHI and HindIII の両方で pAMP を処理すると,
- ampr 遺伝子と複製開始点が含まれる 3755 個の塩基対の断片と,
- 784 個の塩基対の断片が得られ,
- 両断片に付着末端が含まれる。
pKAN
- 4207 個の塩基対
- 単一複製開始点
- 抗生物質 カナマイシン kanamycin( ストレプトマイシンと同じアミノ配糖体に属する抗生物質 )耐性の遺伝子 ( kanr )
- BamHI による単純切断
- HindIII による単純切断
pKAN を BamHI と HindIII の両方で処理すると:
- 2332 個の塩基対の断片
- kanr 遺伝子を含む 1875 個の塩基対の断片 ( ただし,複製開始点は含まれない )
- 両断片に付着末端が含まれる。
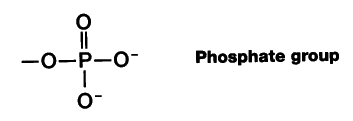
制限酵素で処理した試料をアガロース・ゲルなどで 電気泳動 electrophoresis することによって,これらの断片を視覚的にとらえることができる。直流電を付加すると,負に荷電している リン酸基 をもつために, DNA は陽電極方向へ泳動する。小さい断片ほど,ゲル内を早く移動する。
{kind=link}
いろいろな断片の再結合
2種類の制限酵素を取り除き,DNA リガーゼ が働くような状態にすると,これらのプラスミドの断片は再結合してしまう。
pKAN と pAMP の断片を混合すると,それぞれの分子が再結合する可能性も複数ある。また,場合によっては,つまり2個の複製開始点を含んだり,1個も含まない分子は機能的なプラスミドに戻らないこともあるだろう。
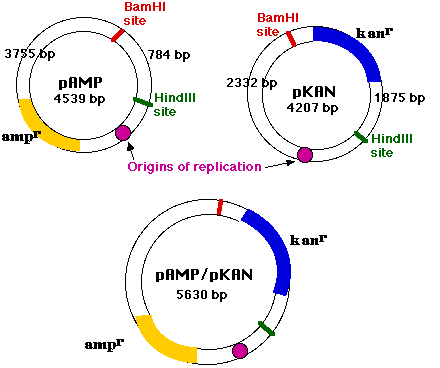
興味深い可能性としては,
- 3755 個の塩基対の pAMP 断片 ( ampr 遺伝子と複製開始点を含む ) と
- 1875 個の塩基対の pKAN 断片 ( kanr 遺伝子を含む ) との結合である。
DNA リガーゼで連結させると,これらの分子は,アンピシリンとカナマイシンの 両方 に耐性を示す機能的なプラスミド( の様 )になる。これが 組み換え DNA 分子の例である。
プラスミドのように機能的な分子として振る舞えるのは複製開始点があるためであるが,これは pAMP に依存するものである。したがって,この場合 pAMP を ベクター と呼ぶ。
大腸菌の遺伝子組み換え
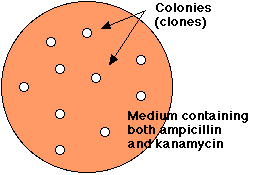
再結合した分子の混合液で大腸菌を処理すると,アンピシリンとカナマイシンの両方が存在しても発育できるコロニーが出現する。
- 大腸菌の混濁液を,再結合した DNA 分子の混合液で処理する。
- アンピシリンとカナマイシンを含む寒天培地に大腸菌混濁液を塗布する。
- 翌日,両方の抗生物質に耐性を示す細胞が,数億にも達する形質転換した細胞のコロニーとして確認できるまで増殖する。
- それぞれのコロニーは遺伝子組み換え細胞の クローン である。
しかし,大腸菌は複数のプラスミドで同時に遺伝子組み換えできるので,形質転換した細胞が組み換えプラスミドをもっているか確認しなければならない。
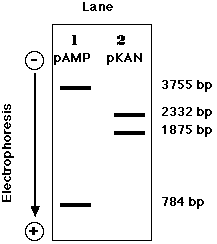
二重の耐性を示すコロニー ( クローン ) のDNA を電気泳動すると,いろいろなことが分かる。
- 組み換えプラスミド だけから耐性を獲得した細胞からのプラスミド DNA は 3755-bp と 1875-bp のバンドのみを示す ( Clone 1, lane 3 ) 。
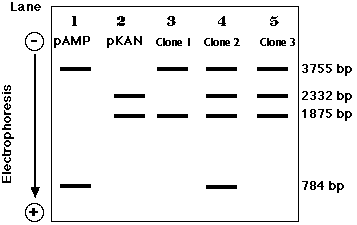
- Clone 2 ( Lane 4 ) は再結合した pAMP と pKAN によって同時に遺伝子が組み換えられた。( この場合,組み換え分子も受け継いでいるかどうかは断定できない )
- Clone 3 ( Lane 5 ) は pKAN と同様に組み換え分子によっても形質転換されている。
遺伝子のクローニング
上述した組み換えベクターはそれ自身他の遺伝子のクローニングのためのツールとして有用である。 今,その カナマイシン耐性遺伝子 ( kanr ) 内に次に単一配列があったと仮定しよう。
5′ GAATTC 3′
3′ CTTAAG 5′
これは,制限酵素 EcoRI で切断でき,付着端をつくることができる。
ここで,たとえばヒト細胞の他の DNA 試料を EcoRI で処理し,同様の付着端が形成されたとしよう。EcoRI 処理プラスミドと DNA リガーゼを混合すると,少数ながらヒトの分子がプラスミドに導入され,これが大腸菌を形質転換するのに利用できる。
しかし,ヒトの DNA をもつプラスミドによって形質転換されたこれらの大腸菌のクローンはどのように検出できるのだろうか?
その鍵は, kanr 遺伝子内にある EcoRI 領域である。したがって,ヒトの DNA がそこに挿入されると遺伝子の機能は破壊される。
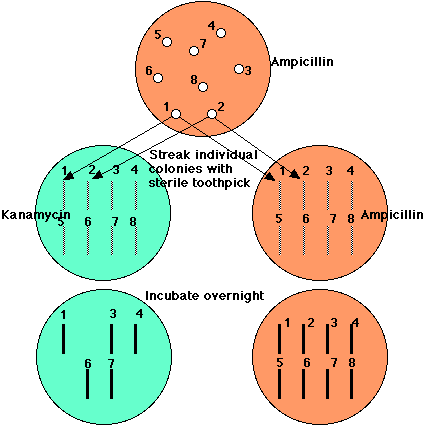
ベクターによって形質転換されたすべての大腸菌は,ヒトDNA をもつかもたないかにかかわらず,アンピシリンの存在下で増殖できる。しかし,ヒト DNA をもつプラスミドによって形質転換された大腸菌はカナマイシンの存在下では増殖できない。
したがって,
- アンピシリンだけを含む寒天に処理した大腸菌の混濁液を塗布する。
- 一晩増殖させる。
- 明瞭なスポットとして同定できるコロニーを滅菌した爪楊枝でカナマイシンを含む寒天に移し替える。
- ( 別のアンピシリン寒天にも同様にする。)
- 一晩培養する。
アンピシリンで増殖するが,カナマイシンで増殖できないこれらのクローンがひとDNA によって形質転換されたものである。 ( ここでは, clones 2, 5, ならびに 8 )
治療に用いられる組み換え DNA 産物
このような手法を用いて,多くのヒト遺伝子が大腸菌や酵母でクローンがつくられている。これはヒトのタンパク質を体外で限りなく生産できることを意味する。ヒトの遺伝子を組み込まれた細胞 ( 大腸菌,酵母,哺乳動物細胞 ) を培養することによって,以下の物質の生産が行われている:
- 糖尿病患者のためのインスリン
- 血友病 A の男性のための血液凝固 第 VIII 因子
- 血友病 B の患者のための血液凝固 第 IX 因子
- ヒト成長ホルモン ( GH )
- 貧血の処理のための エリスロポエチン erythropoietin ( 赤血球生成促進因子 )
- 数種類の インターフェロン
- 数種類の インターロイキン
- 骨髄移植後の骨髄の刺激のための 顆粒球マクロファージコロニー刺激因子 ( GM-CSF )
- (例えば、化学療法の後に)好中球の生産を刺激して,血中に骨髄から造血幹細胞を動員するための顆粒白血球コロニー刺激因子(G-CSF)
- 血栓を溶解する 組織プラスミノゲン活性化因子 (TPA)
- 重症複合型免疫不全 ( SCID ) 治療用の アデノシン・デアミナーゼ ( ADA )
- 副甲状腺ホルモン
- 多くのモノクローナル抗体
- B型肝炎ウイルスに対して予防接種をするためのB型肝炎表面抗原(HBsAg)
- 遺伝性血管浮腫を治療するのに用いられるC1インヒビター(C1INH)
さらに多くの例が開発中である。
March 22, 2020