4.家畜におけるメンデル遺伝
このページの内容
メンデルの人物像や,彼がエンドウを用いて行った実験の詳細についてはこちらを参照してください。
4-1.優劣の法則
メンデルの法則では,個体を構成する細胞に遺伝子が対になって存在し,それぞれ一方の遺伝子が配偶子に分配され,両親から子へ遺伝物質が運搬されることが基本原理となっている(下図)。
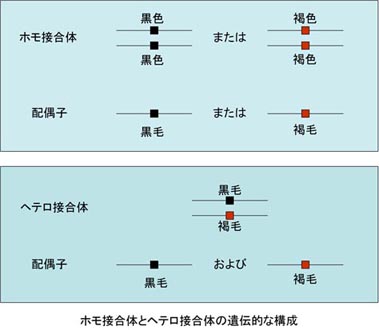
ホモ接合体とヘテロ接合体の遺伝的な構成
遺伝物質上の遺伝子の局在部位を遺伝子座 locus (plural, loci) という。
ある遺伝子座の異なる遺伝子(たとえば黒毛と褐毛の遺伝子)の組み合わせにより,異なる形質が生じる。これらを対立形質と呼ぶ。
一方の遺伝子の効果を隠してしまう対立形質は優性dominantであるといい,隠れてしまう対立形質は劣性recessive であるという。
配偶子の形成過程で,対の遺伝子が雄では精子,雌では卵子に分離する。 これらは個体の遺伝子の半数をもつだけなので,半数体と呼ばれる。
個体の体細胞は2倍体であり,遺伝子がすべて対となって含まれる。
遺伝子座にある異なる対立遺伝子を表わすために,遺伝子の省略記号が使われる。
優性の対立遺伝子は大文字で,劣性の対立遺伝子は小文字で表わす。同一の遺伝子座における対立遺伝子には同じ文字が使われる。
黒毛(B)と褐毛(b)の例では,褐毛の表現型 phenotype をもつ動物は bb と記載される遺伝子型 genotype をもつ。
表現型が黒毛の動物は,検定交配や両親の記録などにより遺伝子型はBB か Bb のどちらかである。B_ という表記法は優性の表現型をもち,遺伝子型が判明していない動物によく使われる。
4-2.分離の法則
〈ウシの毛色の例〉
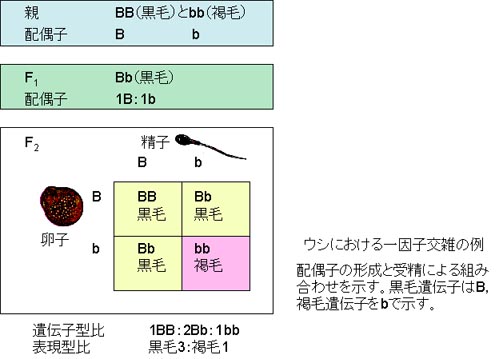
ウシにおける一因子交雑の例
メンデルが次のような動物の交配実験をしたと仮定しよう(左図)。
純粋交配が繰り返し行われている黒毛の動物をつれてきて,褐毛の動物と交配させたとしよう。
1つの形質が異なる純粋種同士の交配は一因子交雑 monohybrid cross という。
両親として選ばれた2つの純粋種は親世代 parental generation という。それらの子畜は雑種第1代 first filial generation,F1 と呼ばれ,この場合,すべて黒毛になることにする。
この場合,一方の毛色が他の毛色を隠してしまうので,黒毛遺伝子を優性であるという。 F1個体同士の交配で雑種第2代 second filial generation F2 が生まれる。
多数のF2個体が得られた場合には,黒毛と褐毛の子畜が3:1 の割合で生まれる。この時,両親で見られた褐毛が再度出現する。
純粋種の褐毛の個体は褐毛遺伝子のホモ接合体(同型接合体)である。ホモ接合体の個体は,この特定の遺伝子対でみると1種類の配偶子だけを生産することになる。
黒毛の場合,どの個体がホモ接合体であるかを検定するためにF2個体を用いて検定交配が必要となる。この場合,褐毛の個体が交配相手として用いられる。その理由は,褐毛遺伝子は,いま検定されようとしている黒毛遺伝子の効果を隠してしまわないためである。
先のF1で見てきたように,黒毛が褐毛に対して優性で,劣性褐毛遺伝子の効果を完全に隠してしまう。
ヘテロ接合体(”ヘテロ”とは異型を意味する)の黒毛の動物は2種類の配偶子を生産する。その半数は褐毛遺伝子をもち,他の半数は黒毛遺伝子をもつ。したがって,ヘテロ接合体は同じ形質の子畜を生むわけではない。
ホモ接合体の黒毛の動物は,その優性遺伝子の効果がすべての子畜に発現するので,検定交配をする必要はない。
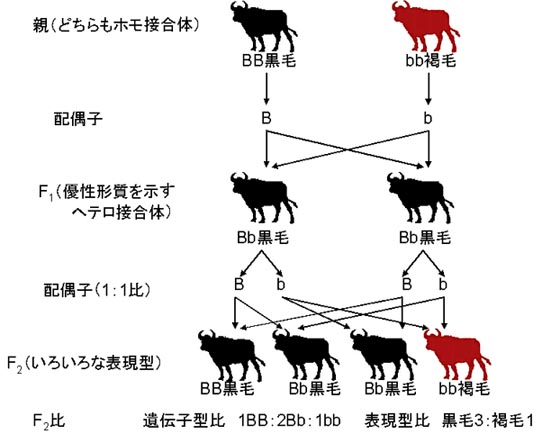
ウシにおける一因子交雑の例
黒毛のF2個体の検定交配では,1/3が黒毛の子畜だけを生産するホモ接合体で,2/3が黒毛と褐毛を1:1の比率で生産するヘテロ接合体である。
ホモ接合体の黒毛と褐毛の数とはほぼ同じなので,全体としてのF2の比率は,
- ホモ接合体の黒毛BB
- ヘテロ接合体の黒毛Bb
- ホモ接合体の褐毛bb
が1:2:1となる。
ヘテロ接合体の検定交配の結果は,遺伝子が対となって個体に伝わるとするメンデルの考え方を支持するものである。そして,これによってヘテロ接合体が2種類の子畜を同数生産することを証明できる。
メンデルは配偶子形成の過程で遺伝子が分離すると推論した。そのため,親はその子畜のそれぞれに各遺伝子対の一方だけを伝えることになる。どの遺伝子が各々の子畜によって受けとられるかは無作為に決まる。
このように,メンデルは彼の最初の実験結果からF1は2つの異なる形質のどちらかをもつ子を生産し,このうち1/2は雑種(すなわち,ヘテロ接合体)となるが,他の1/2は優性または劣性の形質を受け継いだ子が同数形成されることを示した。
4-3.独立の法則
メンデルは次に形質が分離してくる過程における遺伝子の行動を説明しようとした。これはメンデルの独立組み合わせの法則となった。
ここでは,それぞれの形質を支配する遺伝子が独立して分離することについて説明している。すなわち,ひとつの遺伝子座における遺伝子の分離は他の遺伝子座の遺伝子の分離に影響しないのである。
〈ブタの毛色の例〉
豚の品種のハンプシャーは体躯の回りに白帯を生じさせる優性遺伝子をもつ。

二因子交雑のF2個体の比の計算
この遺伝子をWで示し,単色を生じさせるこの対立遺伝子はwで示す。純粋交配をしている褐毛で白帯をもつ豚bbWWを純粋交配している黒毛単色の豚BBwwと交配したとしよう(左図)。
雑種第1代F1の遺伝子型はBbWwとなり,表現型は白帯をもつ黒毛となる。
F2ではどのように発現させるであろうか。これらの形質が独立しているのであれば,それぞれの遺伝子対はメンデルの第2法則に従って分離し,遺伝子型の比が1:2:1となるはずである。左図にF2比を求めるための2つの遺伝子座の比率のかけ算の展開を示す。
F2の表現型の期待値である黒毛3:褐毛1と白帯3:単色1の代数計算が利用できる。
これら2つの比をかけ算することによって,F2の表現型比である黒毛白帯9:黒毛単色3:褐毛白帯3:褐毛単色1が求められる。 すなわち,どちらも優性9:それぞれいずれかが優性3:どちらも劣性1の比として現われる。
2つの形質の対立遺伝子が独立して分離するので,後代は同じ確率でいろいろな組み合わせの対立遺伝子を受け継ぐ。
毛色の遺伝は表現型を支配する異なる遺伝子座の働きを考える際のよい例である。
〈イヌの毛色の例-独立の法則〉
次に示したのはドーベルマン犬における毛色の例である。

ドーベルマン犬における毛色
黒色-タン(ブラック),褐色-タン(レッド),青色(ブルー),ならびに淡黄褐色(イザベラ)
2つの毛色斑が最も一般的で黒色-茶色(タン)と褐色-茶色(タン)である。事実,この’表現型’は毛色と斑紋とに分けて考えることができる。
脚,鼻口部,眉,ならびに胸部にみられる茶色はこの品種に特有の斑紋である。
したがって,ドーベルマンで毛色の遺伝について研究する場合には,主な体躯の毛色が黒色か褐色のいずれかということが問題となる。ただし,この他にまれではあるが2つの毛色,すなわち青色 blue と淡黄褐色 fawn (Isabella) が知られている。
黒色毛と褐色毛の遺伝についてはすでに示した。青色と淡黄褐色は実際はそれぞれ‘希釈された黒毛’と‘希釈された褐毛’ということである。実際の交配結果を見てみると,4つの毛色の遺伝の様式を推論する情報が得られる。
淡黄褐色の個体同士だけで交配すると淡黄褐色のみが生まれるので,劣性ホモ接合体と推測される。褐色毛はすでにbbのホモ接合体であることを示してきた。
したがって,淡黄褐色に影響する対立遺伝子はヘテロ接合体の可能性がある。
毛色を希釈する遺伝子をdで表わし,完全な色を発現する対立遺伝子をDで表わす。
褐 毛=bbD_
淡黄褐色=bbdd
淡黄褐色の子犬を生む褐色毛犬の遺伝子型はbbDdで,純粋の褐色毛犬の遺伝子型はbbDDであろう。褐色毛犬と淡黄褐色犬の交配に関するデータを以下のようにまとめることができる。
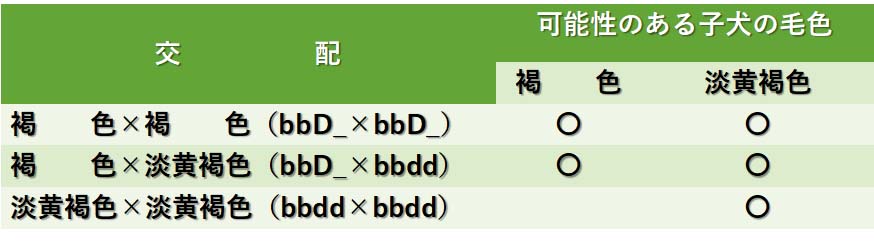
褐色毛犬と淡黄褐色犬の交配によって生まれる可能性のある子犬の毛色
黒色犬と青色犬の交配の例を以下に示す。
黒毛はB_D_であり,青色(黒色の希釈色)はB_ddと仮定する。
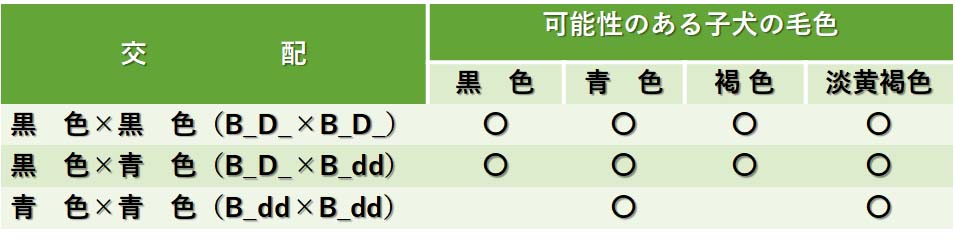
黒色犬と青色犬の交配によって生まれる可能性のある子犬の毛色
これらは交配を重ねると観察される例として示している。繰り返しになるが,特定の交配の結果はそれに関わる遺伝子型に依存して決まってくる。黒色は2つの優性対立遺伝子に依るものなので,いろいろな毛色が生じる可能性を秘めている。
これらの情報をもとに,いろいろな交配の組み合わせを表にまとめると以下のようになる。
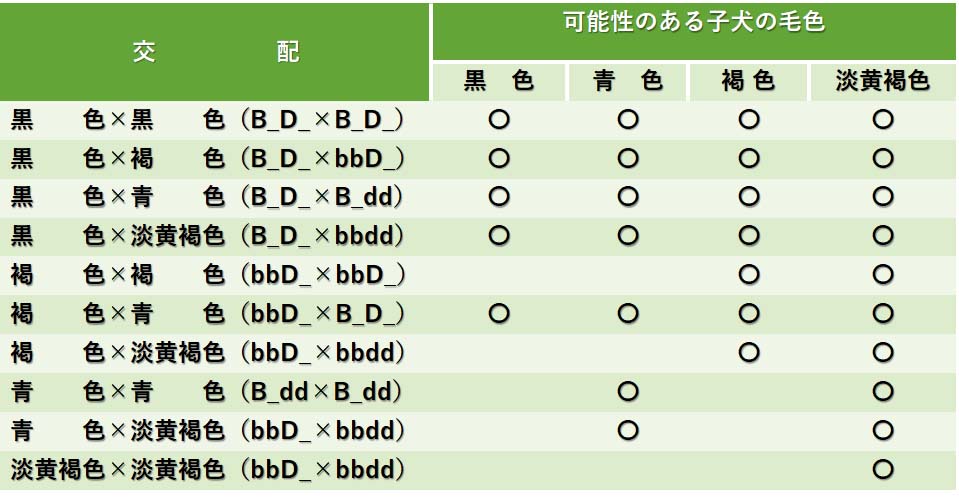
いろいろな毛色の犬の交配によって生まれる可能性のある子犬の毛色
このような表にすると,実際に親の遺伝子型が分らなくてもいろいろな毛色個体間の交配から生じる可能性が予測できる。生まれた子犬を観察すると親の遺伝子型をより正確に推定できるだろう。
純粋交配している黒色ドーベルマン犬を純粋交配している淡黄褐色犬と交配させたとしよう(左図)。すなわち,各系統は数世代を調べても他の毛色の子犬が生まれていないという純粋種である。
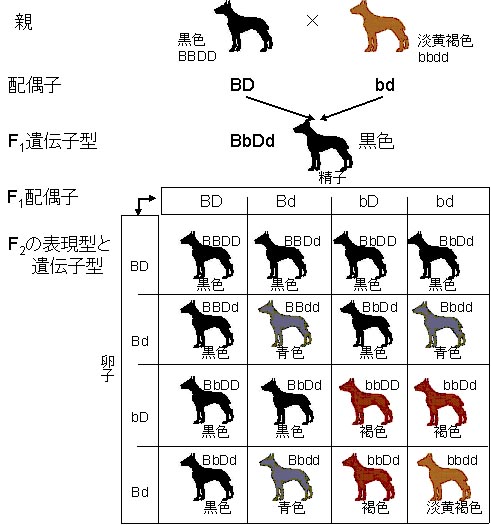
いろいろな毛色のドーベルマンを交配して生まれる可能性のある子犬の毛色
BBDD(黒色)× bbdd(淡黄褐色)
この交配は2種類の遺伝子対が含まれる交配なので,二因子交雑 dihybrid cross と呼ばれる。
F1はすべて遺伝子型がBbDd,表現型は黒色となる。
F1同士を交配させると,各遺伝子対は分離して4種類の配偶子型を生む。 すなわち,BD,Bd,bD,ならびにbdが1:1:1:1の比率で生じる。
1つの表現型BBDDとbbddの交配によって,右図に示すような結果が得られる。
左図に示されたプネットの方形 punnett square (イギリスの遺伝学者R.C. Punnettに因む)は,配偶子の受精による可能性のあるすべての組み合わせを模式的に示している。
F2では9つの遺伝子型があるが,表現型は4つである。多数の子犬が得られる場合には,F2の比はおよそ
黒色9:褐色3:青色3:淡黄褐色1
すなわち,
- 2種類の遺伝子とも優性-9
- いずれか一方が優性-3
- 2種類の遺伝子とも劣性-1
したがって,9:3:3:1となる。
メンデルは後代の情報を集め,彼の数学の知識を駆使してその表現型比からそれぞれの遺伝子の行動を推測し,“交雑種でみられるそれぞれの形質は親世代での異なる形質には影響されず,独立して分離する”と結論した。
4-4.メンデル比の例外
これまで紹介してきた遺伝の例では,一方の対立遺伝子(2本の相同染色体の同一座位にある遺伝子)は他方に対して優性で,そのため表現型で比較するとヘテロ接合体は優性ホモの個体と本質的に識別できない。この現象は完全優性 complete dominance と呼ばれる。
しかし,多くの対立遺伝子は必ずしもはっきりと優性,劣性というようには発現していない。
遺伝子によって作られる特性は,遺伝子同士で,また環境ときわめて密接に影響しあう。これらの相互作用によって,観察される形質に対する遺伝子の働きが変化する場合がある。
4-4-1.複対立遺伝子
〈イヌの毛色の例〉
これまでの議論では,各個体は遺伝子座に一対の遺伝子(各遺伝子座に2つの対立遺伝子)だけをもち,ホモ接合体かヘテロ接合体のいずれかを示すと仮定してきた。
しかし,実際,動物の集団内には多くの対立遺伝子が存在するようである。個体群をいろいろ調べると、特定の座に対して2つ以上の対立遺伝子を見い出すことがある。もし、ある座に対する3つ以上の対立遺伝子が個体群内に存在するならば、その遺伝子座は複対立遺伝子を持つという。
これの複対立遺伝子の例がイヌの毛色と斑紋形成に見られる。
- 茶色(タン)斑と単色は斑紋遺伝子座上の2つの対立遺伝子の作用によるものである。
- 単色(As,こげ茶ないし黒色の均一な色素沈着)は茶色斑(at)に対して優性である。
- この遺伝子座上の3つ目の対立遺伝子がayで,黒色系の色素沈着を制限する作用をもつ。これはatに対しては優性であるが,Asに対しては劣性である。ay遺伝子がホモのイヌでは,セイブル sable と呼ばれる均一な茶色(タン)を生じる。
したがって,優性の順序は,As>ay>atと表わすことができる。 ここでは,最も優性な対立遺伝子を大文字で表記し,上付文字で対立遺伝子を区別している。
複対立遺伝子に対する表現型の交配表を,ドーベルマンの毛色の二因子雑種の例と同じように作成できる。茶色斑の対立遺伝子が他のすべてに対して劣性であるので,そのようなイヌ同士だけで交配した場合に純粋種のはずである。
セイブル犬と茶色斑犬を交配させても,親のどちらも最も優性であるAsをもたないので,単色の子犬が生まれることはない。単色犬(As_)を含めた交配により,各親の2つ目の対立遺伝子に依存していろいろな種類の子犬が生まれる。
たとえば,親が,AsayとAsatであれば,単色(As_)とセイブル(ayat)の子犬が生まれるが,茶色斑の子犬は生まれない。次の表はすべての組み合わせについて示している。
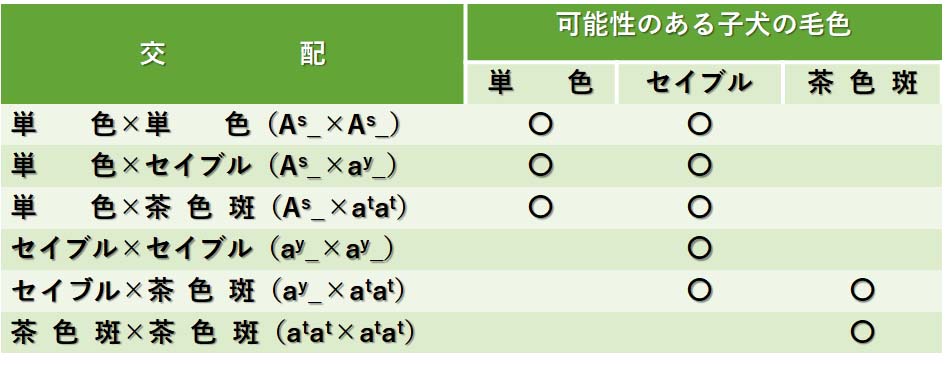
イヌの毛色と斑紋形成に見られる複対立遺伝子の例
ほとんどの毛色の遺伝子座では2つ以上の対立遺伝子が存在するので,これらの各遺伝子座はシリーズ (series)とよばれる。
各遺伝子座(シリーズ)の遺伝子は必ずしもきちんとした優性を示すわけではない。ある対立遺伝子は完全優性であるが,他は共優性で,さらに完全劣性のものもある。
ネコのCシリーズの遺伝子がこの例である。完全色(C)はこのシリーズのすべての遺伝子に対して優性である。
ビルマ(暗赤色)cb遺伝子とシャム(cs)遺伝子はCに対して劣性であるが,お互いに共優性である。cbcsネコはビルマとシャムの中間の毛色斑を示し,トンカTonkaneseと呼ばれる。
目が青く,白毛になる遺伝子(ca)はC,cb,ならびにcsに対して劣性であり,アルビノ(c)は他のすべてに対して劣性である。
したがって,ネコの毛色遺伝子の優性の関係は,
C > (cb=cs) > ca > c となる。
| ヒトのABO式血液型も複対立遺伝子の例であリ,共優性の良い例でもある。 |
4-4-2.遺伝子間の相互作用
不完全優性
1つの遺伝子座にある対立遺伝子が他方の対立遺伝子の発現を完全には阻止しないとき,その対立遺伝子は不完全優性 incomplete dominance を示すという。
たとえば,白い花をつける純系の植物と赤い花をつける純系の植物を交雑させると,ピンクの花だけがF1世代に出てくる。
〈鶏の羽色の例-不完全優性〉
鶏における羽色も不完全優性のよい例である。純系の黒色鶏と純系の白色鶏を交雑させると,アンダルシアン・ブルーと呼ばれる灰色だけのF1鶏が生まれる。
ヘテロ接合体である灰色鶏同士の交雑では,子世代において2つの対立遺伝子は分離して,黒色鶏:灰色鶏:白色鶏が1:2:1の比率で生まれる。
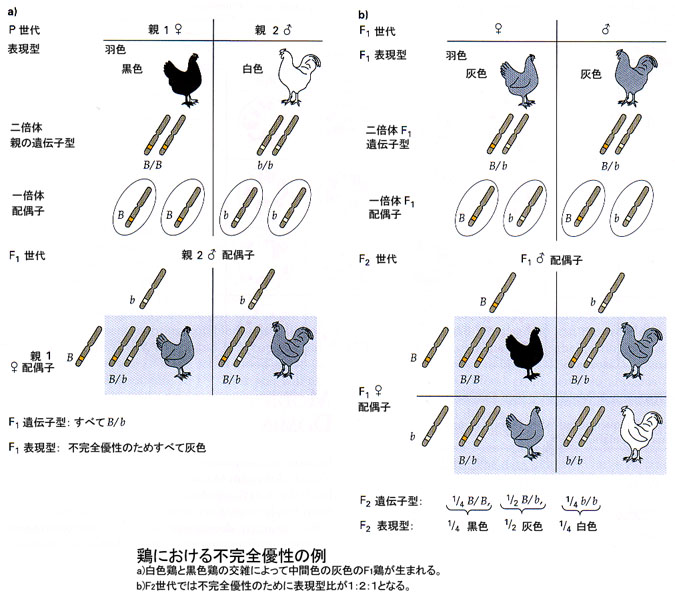
ニワトリにおける羽色の遺伝(不完全優性の例)
共優性
ある遺伝子座における対立遺伝子が同等に発現したとすると,それらは共優性 codominance を示すという。
〔家畜における例〕
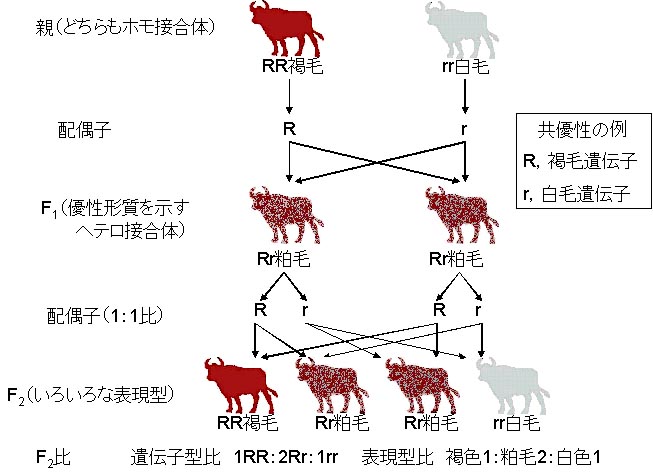
肉牛ショートホーンの褐毛と白毛の個体を交配すると粕毛の個体が生まれる共優性の例(ウシの毛色)
メンデルが牛のショートホーンの毛色変異を知っていたら,一時的にも融合遺伝理論を信じてしまったかも知れない。
純系の褐毛ショートホーンと純系の白色牛を交配させると,どちらの親にも似ていないF1が生まれる。このF1は粕毛(かすげ)と呼ばれ,褐色毛と白色毛が混在して生えている状態になる。
しかし,F2をさらに調査すると,メンデルの法則に合致していることが分る。F2では,褐毛1:粕毛2:白毛1の割合で出現し,これはむしろ完全優性ではない状態を示している。
ヘテロ接合体はどちらのホモ接合体とも異なる特有の表現型を示す。
褐毛,粕毛,ならびに白毛牛のいろいろな交配の組み合わせから生じる子牛の毛色を以下に示す。
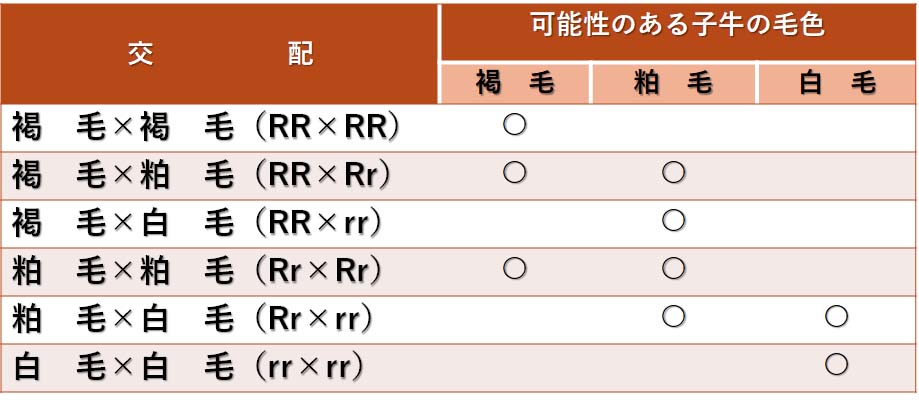
肉牛ショートホーンにおけるいろいろな毛色の個体の交配により生まれる可能性のある子牛の毛色
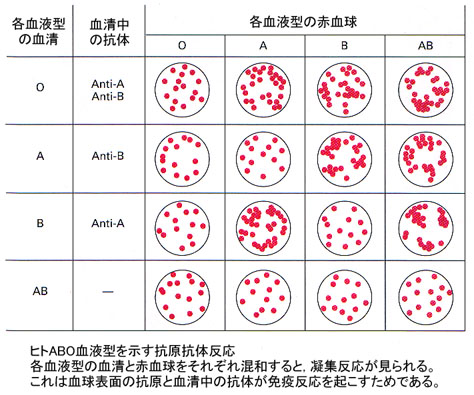
ヒトABO式血液型
ヒトにおけるABO式血液型を決定するAとBという対立遺伝子は共優性を示す対立遺伝子である。すなわち,Aという対立遺伝子とBという対立遺伝子の両方を持っている人の血液型はAB型である。
| ヒトの血液型の詳細についてはこちらを参照してください |
| ABO式血液型についてはこちらも参照してください |
超優性
ヘテロ接合体(Aa)がAAやaaといったホモ接合体よりもずっと強くその表現型を発現するとき,遺伝子間の相互作用は超優性 overdominance である。
この現象は雑種強勢とも呼ばれ,動植物の育種にとって重要な役割を果たしている。
異なる遺伝子座位上にある遺伝子の相互作用
これまで紹介してきた二因子交雑の例では,それぞれの遺伝子座に由来する表現型は独立して遺伝するものであった。しかし。2つの遺伝子座が1つの形質の発現に関与する例があり,それらの遺伝子の相互作用によってまったく新しい表現型が生じることがある。
〈鶏の鶏冠の例〉
バラ冠遺伝子(R)とマメ冠遺伝子(P)の相互作用によって,まったく新しいクルミ冠(R/- P/-)が生じる。

鶏のトサカの遺伝
上位性(エピスタシス)
1本の染色体上の1遺伝子座にある対立遺伝子が,離れたところにある対立遺伝子の発現に影響する,すなわち隔たった遺伝子座にある遺伝子間の作用を上位性 epistasis という。
| 上位性と優性を混同しないこと |
|---|
| 優性とは,同一の遺伝子座において一方の対立遺伝子が他方の対立遺伝子の発現を抑えることである。 |

毛色を支配する遺伝子
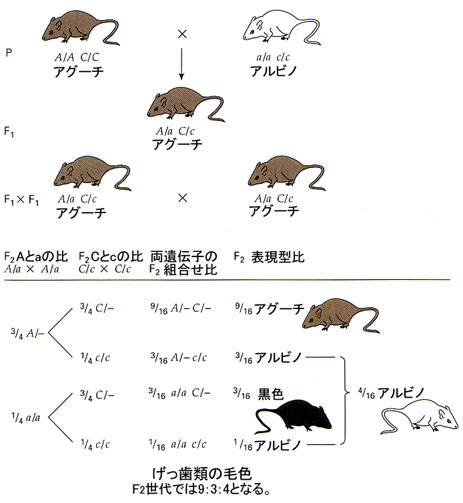
げっ歯類の毛色の遺伝(上位性遺伝子の例)
致死遺伝子
致死遺伝子とは,それが発現した場合に出生前後に胎子の死を引き起こすもので,半致死遺伝子とは発現すれば生存はできるが,重い病気または障害を起こすものである。
〈馬における致死遺伝子の例〉
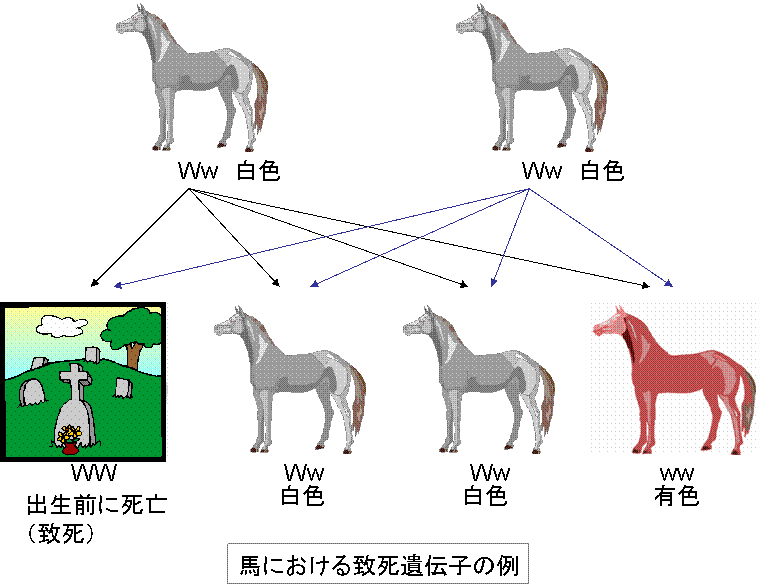
馬における致死遺伝子の例
早期胚死亡の例が馬における優性白毛に関連してみられる。
この遺伝子座の2つの対立遺伝子が白色となるか,あるいは有色となるかを決める。
白色被毛は他の遺伝子座の遺伝子の影響を受ける。優性白色の表現型をもつ馬は彼らの間だけで交配させると白色子馬と有色子馬が2:1の割合で生まれる。
なぜ白色馬と有色馬の比率が3:1ではなく,2:1なのであろうか。結論は期待される白色馬の1/3が後代として生まれてこないということである。
4-4-3.遺伝子と環境間との相互作用
遺伝子は他の遺伝子の発現によって影響を受けるばかりでなく,
- 体内環境
- 年齢
- 性(ホルモン)
ヒトの禿,鶏の羽装,ドーセットホーンの角
- 体外環境
- 温度
ヒマヤラ・ウサギやシャムネコの毛色 - 栄養
- 光線
- 化学物質
- 微生物の感染
- 温度
発現度変異
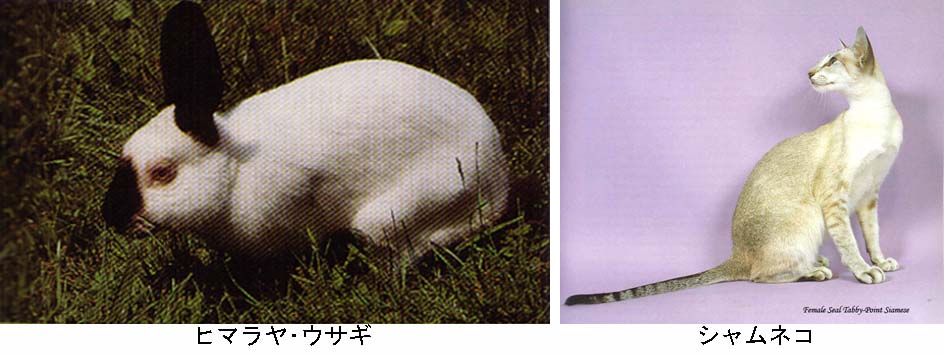
発現度変異の例
ヒマラヤ・ウサギ(左)やシャムネコ(右)の毛色。

気温によって変化するヒマラヤ・ウサギの毛色パターン
生物個体のそれぞれの細胞に存在している1対の対立遺伝子が互いに異なった発現をする場合,それらは発現度変異 variable expressivity を示しているという。
よく知られている例として,ヒマラヤ・ウサギやシャムネコの毛色がある(写真参照)。これらの動物の尾の先端,耳,四肢は黒ずんだ色をしているが,他のところの毛の色は一様に白っぽい色をしている。
皮膚のすべての細胞は毛色に関して同じ対立遺伝子を持っているが,発育過程における体のそれぞれの部分の体温が毛の発色に影響する例である。
尾の先端,鼻先,耳,四肢は体の主な部分よりも概して体温が低いので,それらの場所において発生した毛はより黒ずむことになる。
これらの遺伝子は温度感受性型のチロシナーゼ tyrosinase の産生に関与し,体の端は他の部分に比べ温度が低いので,チロシナーゼが体の端に近いほど効果的となる。
発現度変異は禿とか病気のかかりやすさに至るまで,人の多くの遺伝形質に影響を及ぼしている。
遺伝病での発現度変異の現れ方
同一家系の(同じ変異アリルを継承している)患者でも,重篤度や発症部位が異なる場合がある。
《例》 マルファン症候群を引き起こすfibrillin 遺伝子の変異はある家系において眼の異常を引き起こし、他の家系では心臓の異常を引き起こす。極端な例では,不完全浸透(後述)を示す場合もある。
例:早発性禿頭
ネコのメインクーンの遺伝性の疾患
{kind=link}
{kind=link}
多面作用
1つの遺伝子が表現型に関して多種多様の作用をする現象を多面作用 pleiotropy という。
メンデルが研究した遺伝子は1つの特徴(すなわち豆の色,形など)のみに影響を及ぼすのに対して,動物の多くの遺伝子は,1つの表現型のいくつかの面に影響を及ぼすことがある。
多面作用は,病気の原因となる欠陥のある対立遺伝子を保留している人にとりわけ顕著である。
例:馬の毛色と致死性(前述)
ヤギにおける角性と間性
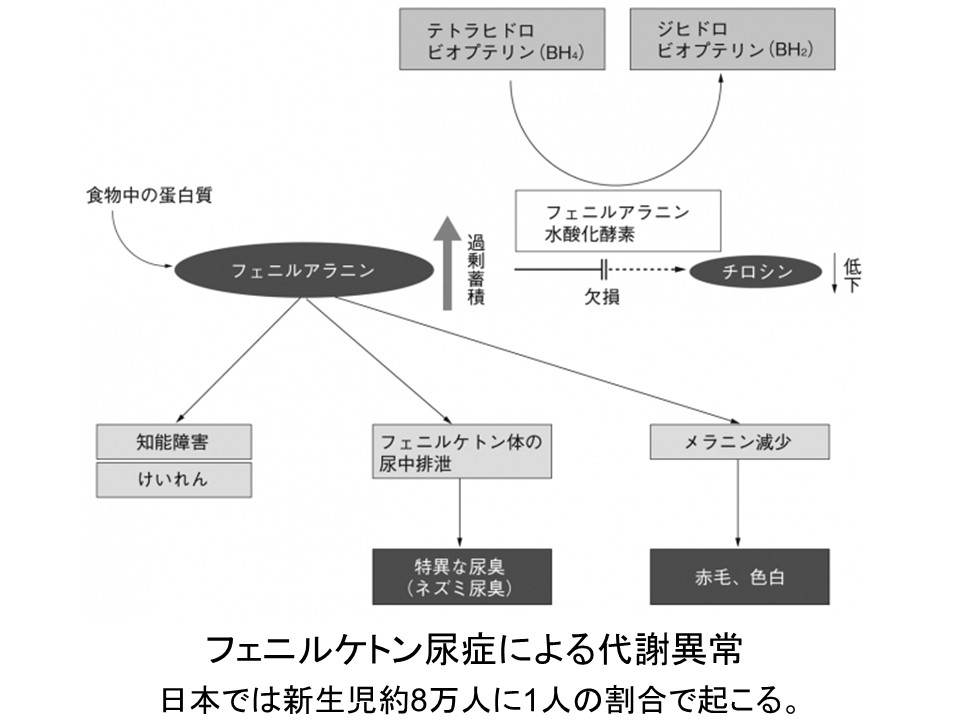
ヒトにおけるフェニルケトン尿症(Phenylketonuria, PKU)
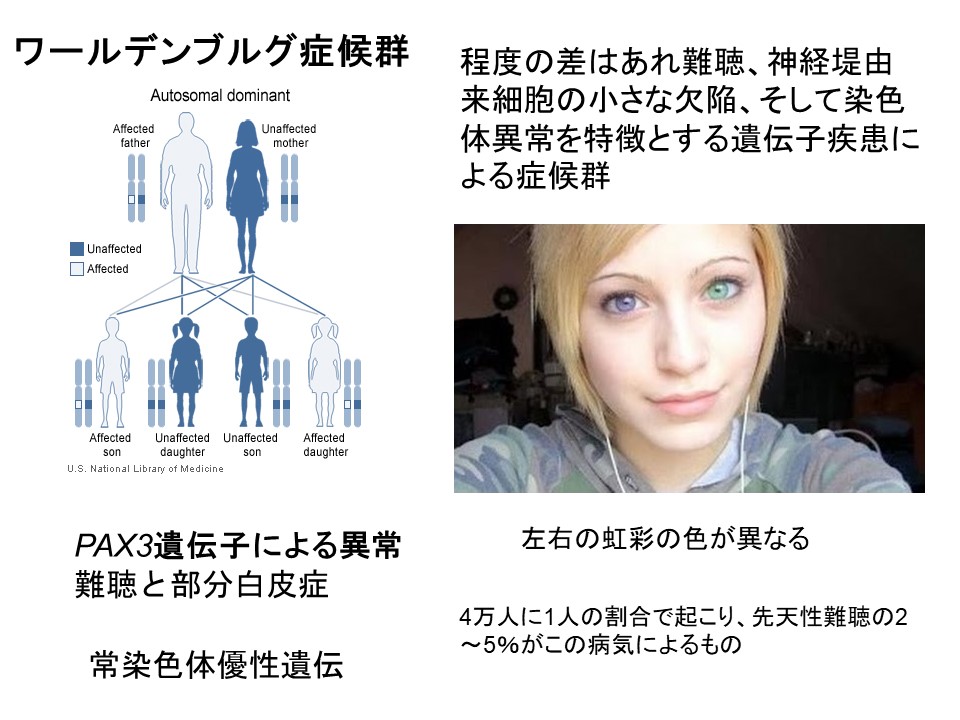
Pax3遺伝子:ワールデンブルグ症候群I型の難聴と部分白皮症
RET遺伝子:ヒルシュスプルング病と多発性内分泌腫瘍症
{kind=link}
{kind=link}
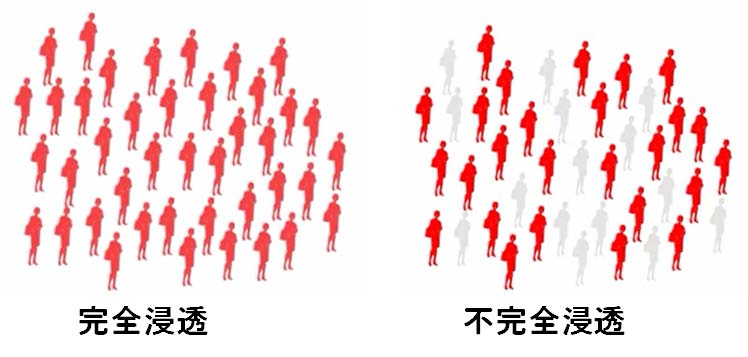
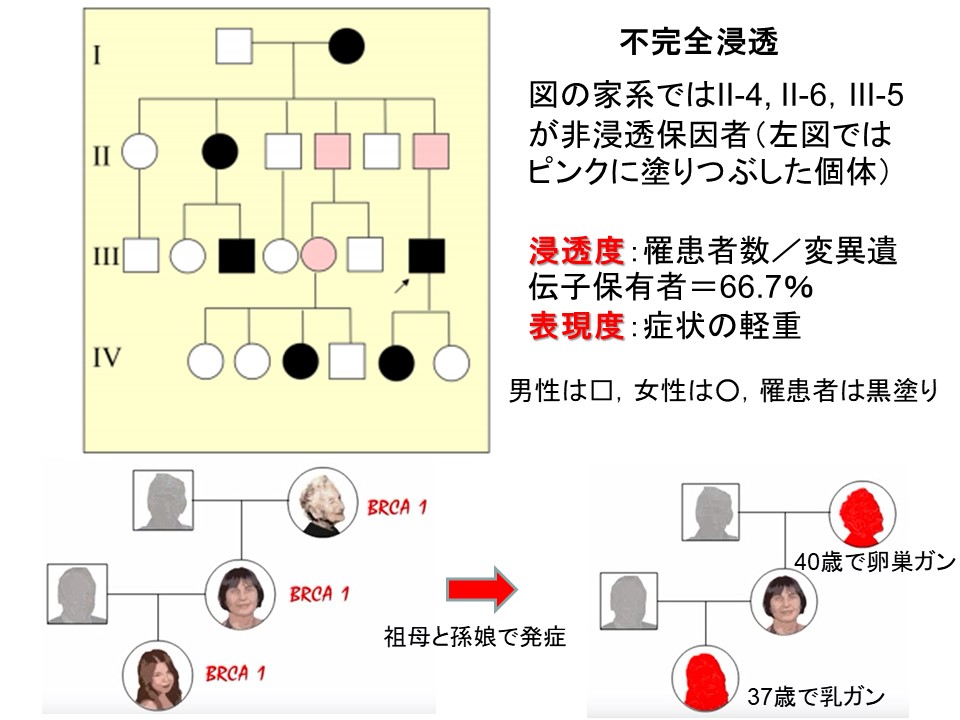
不完全浸透
病気の原因となる遺伝子(優性遺伝子で1コピー,劣性遺伝子なら2コピー)が個体に存在しても必ずしも変化を引き起こさなかったり,不完全な変化しか起こさない場合がある。
このように,普通は特別な表現型を生じさせる遺伝子が発現しない場合を不完全浸透 incomplete penetrance といい,そしてその度合いを不完全浸透度という。
必ずしも発病しなかったり,不完全な変化しか起こさない場合を不完全浸透という。
ある遺伝子が,異なる個体において同じく発現しないのは,遺伝的かつ環境的影響が組み合わさった結果である。
例:ヒトや家禽における多指症
BRCA1変異遺伝子によるガン
{kind=link}
| 最初に戻る |
| メニューのページへ戻る |
March 02, 2020