| このページの内容 |
5.染色体とその動態
家畜の育種は染色体レベルで操作するわけではないが,染色体の構造と挙動についての知識が必要である。
5-1.家畜の染色体
分裂途中の細胞を染色して顕微鏡でみると,それぞれの核の染色体が観察される。
しかし,染色体標本を確実に作製するのはなかなか難しい技術である。種によって適切な条件が異なるため,多くの種で染色体が研究されるのにはかなりの時間が必要であった。
1956年にはヒトの正確な染色体数が決定されている(Tjio and Levan, 1956)。それぞれの種の染色体構成を核型 Karyotype という。
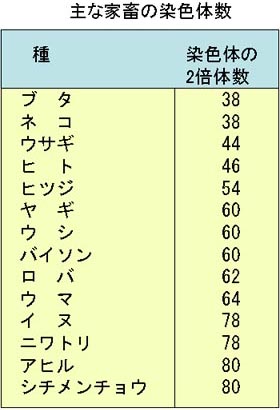
例として,ブタ(オス)とニワトリ(オス)の核型(図)と,主な家畜の染色体数(表)を示す。
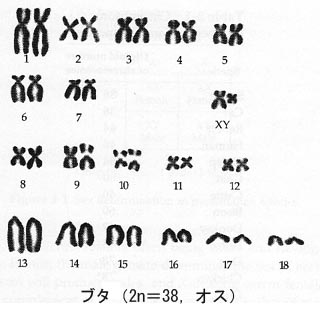
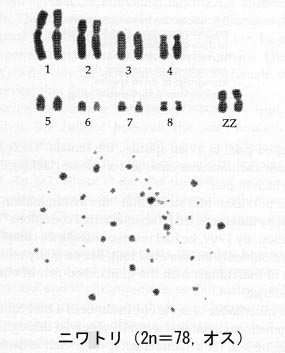
5-2.染色体と性の決定
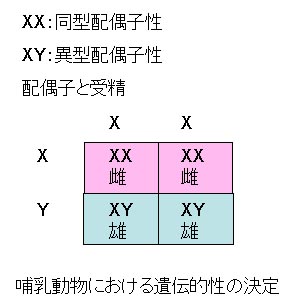 いろいろな種で核型が研究されるにつれ,すべての染色体が対で存在するという原則の例外があることがわかってきた。
いろいろな種で核型が研究されるにつれ,すべての染色体が対で存在するという原則の例外があることがわかってきた。
すなわち,一方の性ではすべての染色体が対を示すが,もう一方の性では一対の染色体が一致しない。
哺乳動物ではオスで,鳥類ではメスで一致しない対が見られる。 性の遺伝は染色体によって制御される形質の一例である。右図に示すように,ウシのような哺乳動物では,すべての卵子は1本のX染色体をもつので,雌は同型配偶子性 homogametic とよばれる。
雄は異型配偶子性 heterogametic で,精子の半分はX染色体をもち,他の半分はY染色体をもつ。 Y染色体はX染色体に比べ小さく,異なった形状を示す。
したがって,Y染色体をもつ精子により雄となり,X染色体をもつ精子により雌となる。
哺乳動物では,Y染色体をもたない個体が雌となる。
Y染色体の存在によって雄として発生するが,ヒトにおける精巣性女性化症候群の遺伝子によって正常に発生しないことがある。
この遺伝子は遺伝的には男性(XY)の個体に女性の外部生殖器を発達させる。
哺乳動物と違って鳥類では,雄は同型配偶子性で雌が異型配偶子性である。鳥における性染色体はZ染色体とW染色体として表わされ,雄がZZで雌がZWである。
| 性の分化についてはこちらを参照してください |
5-3.染色体の形態
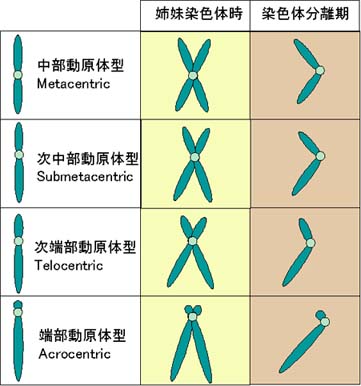 染色体の形態は種によって大きく異なる。
染色体の形態は種によって大きく異なる。
核型は分裂期の染色体像を示すもので,染色体が複製され,それぞれ分離する前(いわゆる姉妹染色体)の時点で細胞周期を停止させる。
姉妹染色体は動原体でお互いに結合している。
- 中 部動原体型 metacentric-中央に動原体があるXのような染色体
- 次中部動原体型 submetacentric-中央からやや外れている染色体
- 次端部動原体型 telocentric-さらに端によっている染色体
- 端 部動原体型 acrocentric-動原体が染色体のほとんど末端にある染色体
(動原体の反対側にかなり小さな染色体部分が存在する)
5-4.有糸分裂
すべての体細胞は有糸分裂によって増殖するが,生殖細胞は減数分裂によって染色体を複製する。
有糸分裂は細胞の増殖と分裂という細胞周期 cell cycle 全体の中のごく一部である。
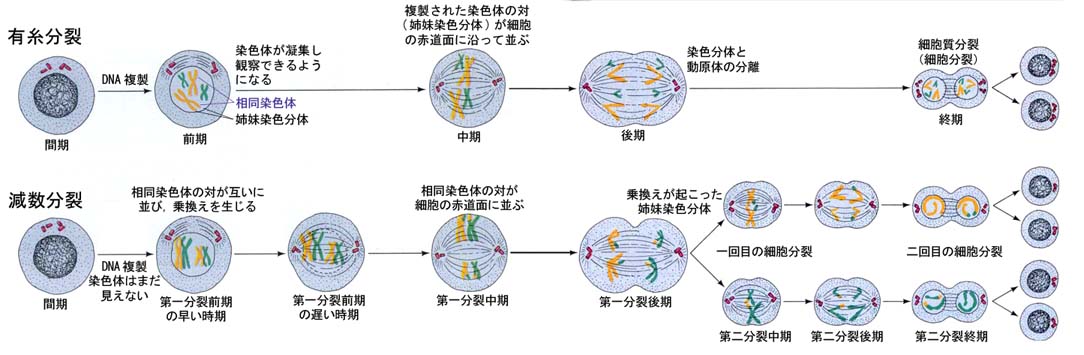
有糸分裂と減数分裂とを比較した模式図を下に示す。
細胞周期のうち最も長い期間である間期では,染色体は拡散して顕微鏡下でも観察できない。
細胞分裂が進行すると染色体が凝集して明瞭に観察できるようになる。それまでに,染色体は複製され一対の姉妹染色体となる。
染色体は細胞の赤道面に配列し,動原体が分離して,姉妹染色体のそれぞれが両極に移動するようになる。
このステージでは,“紡錘体”が細胞内に認められ,細胞の両極にある中心小体から動原体に紡錘糸が伸張している。
核膜はこの時期には消失している。紡錘糸が姉妹染色体を赤道面から両極に引き離し,紡錘糸が消失すると2つの核膜が形成され,2個の娘細胞が生じる。
| 有糸分裂の詳細についてはこちらを参照してください |
5-5.減数分裂-遺伝的多様性の源
減数分裂は,有糸分裂と次の4点で明らかに異なっている。
- 減数分裂は生殖細胞で行われ,体細胞では行われない。
- 減数分裂は配偶子に含まれる染色体の本数を二倍体の本数(2n)から一倍体の本数(n)に減少させる。
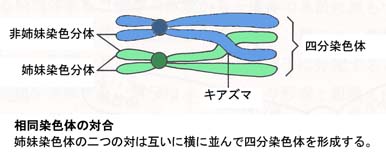
- 相同染色体が減数分裂の最中にぴたりと並び対になる。これらの4本の染色分体は四分染色体と呼ばれる(上図,第一減数分裂中期)。
- 減数分裂の際に,対になった相同染色体同士で乗換えが起こり,遺伝情報の新しい組み合わせが生じる。
| 減数分裂の詳細についてはこちらを参照してください |
雌における減数分裂(卵子形成)と雄における減数分裂(精子形成)の特徴は以下のとおりである。
- 雄では精巣における減数分裂の結果,4個の精子が形成される。
- 雌では,1個の細胞が生殖細胞,すなわち卵子であり,他の3個は生殖細胞としての機能をもたない。
- 雌における第1減数分裂は第2卵母細胞が排卵される時に完了する。この分裂における小型の細胞は第1極体と呼ばれ,発生に関与しない。
第2減数分裂は精子が卵子に侵入するまで起こらない。この分裂の後に,精子と卵子の染色体が合一して,接合体が発生し始める。 減数分裂により,父方由来と母方由来の相同染色体がランダムに分配され,遺伝的に多様な生殖細胞が形成される。
たとえばウシの場合,30対,計60本の染色体をもつので,その組み合わせは230通りとなる。
さらに,減数分裂によって染色体の組換えが起こる。
たとえば,ウシで1本染色体に1ヵ所ずつ組換えが起こったとすると,その組み合わせは260種類となる。
このように考えると,同じ遺伝子の組み合わせをもった配偶子はほとんどあり得ないことになる。
| ウシの場合 | 230=1,073,741,824≒109260=1,152,921,504,606,846,976≒11×1017 |
| ヒトの場合 | 223=8,388,608≒8.4×106246=70,368,744,177,664≒70×1012 |
| 減数分裂時に,遺伝子と染色体がどのように動くのかを示したページは こちらを参照のこと。 |
受精によって倍数性が復元する
 減数分裂は生物の生殖に関して3つの必須な働きをしている。
減数分裂は生物の生殖に関して3つの必須な働きをしている。
- 減数分裂により配偶子に含まれる染色体数を二倍体の本数から一倍体の本数に減らすことによって,世代から世代にわたって,二倍体の染色体数を維持する。
- 減数分裂は染色体を無作為にぶんりし,互いに影響を及ぼしあうことなく配偶子に分配することによって,続く世代に遺伝的な多様性を保障する。
- キアズマの形成と組み替えによって,減数分裂は双方の親の遺伝子の組み合わせとは異なる新しい組み合わせを持った配偶子を生じさせる。そのような遺伝的な差異によって,子孫がある特定の環境下でもっと最もうまく生き延び,繁殖することが可能となる。Darwinが主張したように進化の基礎となるのかもしれない。
キアズマと交叉
細胞分裂中に,同一の染色体上の遺伝子座間で交叉が起こる。
第1減数分裂の複糸期(染色四分体が対合している)に,長軸に沿って縦裂し計4本の染色分体 chromatid となるが,この4本の染色分体の間で互いに相手を交換する部位がみられる。この部位をキアズマという。キアズマは交叉を意味するギリシア語に由来する。
5-6.染色体の異常
5-6-1.染色体の不分離
遺伝情報を忠実に伝達するために,減数分裂は極めて正確な機構でなければならない。しかし,配偶子形成の過程で過誤が起こり,その影響が個体に現われることがある。
有糸分裂でもエラーは起こるが,この場合には体の限られた細胞にのみ遺伝的な影響が現われる。
まず,染色体全体に関わる減数分裂のエラーについて見てみよう。
倍数性 polyploidy - 染色体の完全な組の過剰をいう。
2倍体の配偶子と正常半数体の配偶子が融合して接合体が生じると3倍体となり,3セットの染色体をもつ個体が生じる。父親と母親の染色体セットが融合すると4倍体となる。
植物では,化学的に誘起した倍数性作出が新しい異品種や雑種の開発の有用技術となっている。しかし,動物では倍数性は致死的で,胚発生途上で死滅してしまう。したがって,倍数性は植物で活用されてはいるが,動物の育種には利用できない。
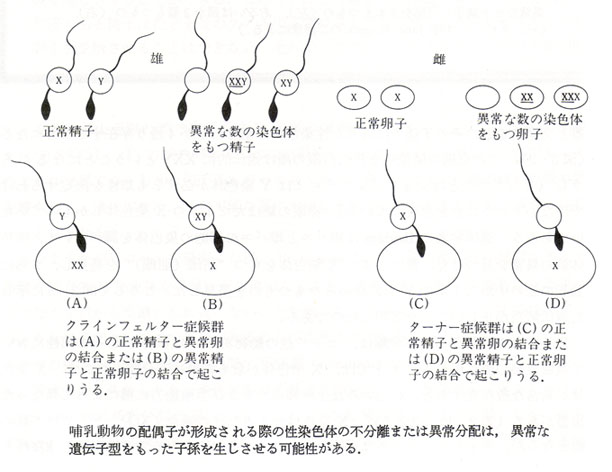
異数性 aneuploidy - 動物では,完全な染色体セットが複製されないことがあり,1~数本の染色体の過不足の状態をいう。
余剰の染色体がある場合を多染色体性 polysomy という。減数分裂中に1本の染色体の分離が遅延したりすると,本来の娘細胞に収まらないことがある。この場合減数分裂の不均等分離が起こる。
この染色体を欠如した生殖細胞との受精によって一染色体性 monosomic となる。逆に,余剰の染色体をもつ生殖細胞が受精すると三染色体性 trisomic となる。
多くの遺伝情報が欠損するのでモノソミーの場合は生存が困難になる。 ただし,性染色体はこの例外である。性染色体の多染色体性の他に,小型の染色体のトリソミーは生存可能なことがあるが,先天性異常を現す。
右図には性染色体の不分離から生じる異常の例を示す。
5-6-2.性染色体の不均衡の補正
余剰のX染色体をもつ個体は,大型の染色体が余剰の場合生存しないという原則の例外である。 X染色体は大型であるが,不均衡が起こった個体も生存する(たいてい不妊である)。
大型の常染色体が余剰にあると生存しないのに,なぜ余剰のX染色体をもっていても個体が生存するのかという理由は性染色体の正常な機能と関連がある。
| 異型配偶子の性では性染色体の遺伝情報は正常な場合でも不均衡である。すなわち,X染色体は大型で,多くの形質に影響する遺伝子座ともつが,Y染色体は小型で遺伝子座も少ない。同型配偶子の性ではその性染色体の機能的遺伝子は異型配偶子のものに比べほぼ倍である。 |
X染色体の不活化が起こり,これが余剰のX染色体があっても生存できる理由である。この現象は遺伝子量補償(補正)として知られている。
したがって,X染色体の総体的な影響は同系配偶子の性も異型配偶子型の性と同様である。
X染色体不活性化の典型的な例が三毛ネコの毛色である。黒毛と褐毛はX染色体上の遺伝子座にある遺伝子によって支配される。体のうち黒毛となる部分は褐毛遺伝子をもつX染色体が不活化された細胞から生じており,褐毛となる部分は黒毛遺伝子をもつX染色体が不活化され多細胞から生じている。
三毛ネコの白斑は白色毛と有色毛(黒色または褐色)の明瞭な領域を決める遺伝子をもつ。斑紋遺伝子は,発生の初期に不活化されたいずれかのX染色体をもつ細胞と同じ起源の細胞から生じる。これは,X染色体の不活化が遅く起こり,黒毛と褐毛が拡散したようなパターンを形成する白い部分のない黒と赤のべっこう型(トータシェル)の場合と大きく異なる。
雄の三毛ネコは稀でXXYをもつ染色体不分離の例となる。この場合,余剰のX染色体が不活性化するため生存は可能であるが,不妊である。
5-6-3.突然変異
染色体を化学的に明らかにしようと多くの研究者が挑み,ついに1953年にJames WatosonとFrancis Crickが染色体がDNAの二重らせんからなることを明らかにした。
DNAの塩基は3個ずつでコドンを形成し,それぞれがアミノ酸や複製開始または終了コドンとなっている。
したがって,DNAは数種類のRNAの鋳型として働く。
mRNAは核から離れ,細胞のrRNAとtRNAと協同して,アミノ酸配列を決めるための鋳型となる。このように,塩基配列は遺伝子そのものである。
遺伝子産物はアミノ酸が配列したペプチド鎖である。RNAテンプレートやリボソームから離れると,ペプチド分子は三次元構造を示すようになり(その構造は構成中のアミノ酸同士の化学的性状によって異なる),他のペプチド鎖と集合することもある。この三次元の構造によって,細胞における遺伝子産物(タンパク質)の活性が決まる。
このようなDNAの構造のために遺伝情報の複製は正確であり,遺伝の機構として極めて重要である。しかし,複製時にエラーが起こることがある。そのようなエラーが複製され集団内に遺伝的な差異を生じることがある。選抜を行う時,このような変異をもった育種集団が必要である。
細胞内の遺伝情報の変化を突然変異と呼ぶ。単一の遺伝子座の突然変異率は極めて低いが,突然変異はすべての体細胞に起こり得る。
点突然変異が起こると,細胞分裂で複製され,すべての子供に伝達される。表現型が変化して,突然変異が起こったことが分る場合もある。外見に現われない,代謝レベルの変異の場合には生化学的な検査が必要となる。
| 突然変異の詳細についてはこちらを参照してください |
| 遺伝子翻訳の詳細についてはこちらを参照してください |
5-6-4.染色体の構造異常
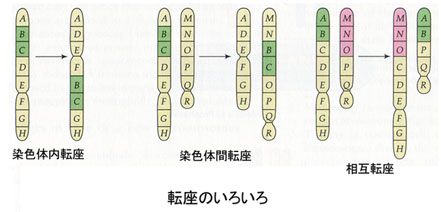
染色体の構造の異常にも種々のものがあるが,ヒトの場合,最もよくみられるのは,減数分裂に際して,一度断裂した染色体の一部がまちがった部分についてしまう転座である。
- 相互転座 reciprocal translocation
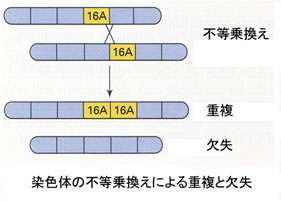
異なった2本の染色体の一部が切断し,互いの切断片が交換して再結合したもの。転座が起こった動物では受胎率の低下が知られている。 - 重複 duplication または欠失 deletion
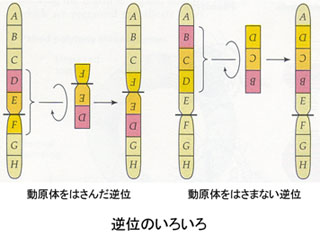
ある染色体で切断が起こり,相同染色体に切断片が結合すると遺伝情報の重複が起こり,切断が起こった染色体では欠失となる。重複と欠失の影響は様々であり,当該部分に含まれる遺伝情報の量や内容によって表現型への影響の程度が違ってくる。 - 逆位 inversion
同じ染色体で2ヵ所以上に切断が起こり,切断片が反転したもの。減数分裂中の対合がうまくいかないので,受胎率の低下が予想される。
| 染色体の構造異常の詳細についてはこちらを参照してください |
| 最初に戻る |
| メニューのページへ戻る |
February 03, 2020