| このページの内容 |
細胞の興奮
刺激を受け興奮した細胞には微弱な電流が検出される。
| 次のページも参照のこと |
- 筋肉細胞 と
- 神経細胞 ( ニューロン )が刺激に対し反応する。
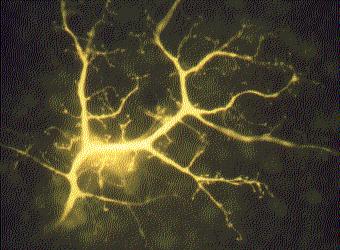
上の写真はウサギ網膜内の 1 本の ニューロン を示す。細胞に蛍光色素が注入され,すべての側枝が観察できる。各側枝先端の膨らみは他のニューロンとの シナプス を形成している。
- ニューロンにおける電流が,シグナルを急速に伝達する。
- 筋肉内の電流が収縮を開始する。 [ 詳細についてはこちら ]
静止電位
すべての細胞は( 刺激を受けていない状態で )静止電位をもつ。細胞膜 を挟んで,細胞の内部は外部に比べ負の電位をもつ。静止電位は変動するが,刺激を受けた細胞では約 -70 ミリボルト (mv) となる。
静止電位には,次のの 2 つの機能がある:
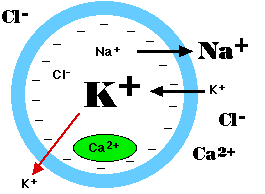
- ナトリウム/カリウム ATPase – ナトリウム・イオン ( Na+ ) 3 個を細胞外へ排出する度に,カリウム・イオン ( K+ ) 2 個を細胞に入れる。したがって,活発に作動すると細胞内の正の荷電が減少する。
- 細胞膜のカリウム伝達通路から細胞外へのカリウム・イオン ( K+ ) の促進拡散が起こるようである(図の赤矢印)。
細胞内のイオンの動き
ナトリウム/カリウム ATPase によって:
塩化物イオン (Cl–) とカルシウム・イオン (Ca2+) 濃度も細胞外で高く維持される。ただし,細胞内にある小胞内に蓄積されている Ca2+ は例外である( 図中の緑色 )。 |
 |
脱分極
外部刺激によって,細胞膜間の荷電が低下することがある:
- 機械的な刺激 ( たとえば,引き伸ばし, 音波 ) によって,ナトリウム伝達通路( 機械的開閉型 )が作動する。
- ある神経伝達物質 ( たとえば,アセチルコリン acetylcholine ) はナトリウム伝達通路( リガンド開閉型 )を開口させる。
それぞれの場合で,細胞にナトリウムが促進拡散すると,細胞のその領域で静止電位が低下し, 興奮性シナプス後電位 excitatory postsynaptic potential ( EPSP ) が形成される。
静止電位が閾値より低下すると( 哺乳動物のニューロンでは約 -50 mv ),細胞内で活動電位が生じる。
活動電位
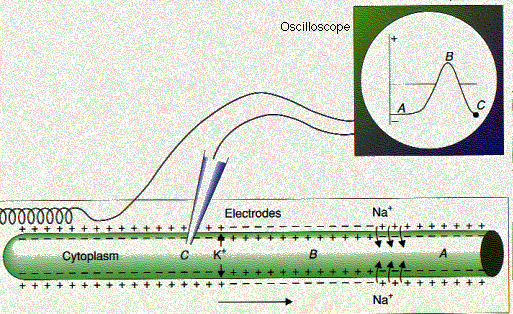 |
| 神経インパルス。 休息中のニューロンでは,軸索内部が外部に比べ負の電位を有している ( A )。活動電位が起こると ( B ), 極性が逆になる。そして, K+ イオンの流出によって,正常な極性が速やかに回復する ( C )。 |
細胞上の一部の脱分極が閾値電位に達すると,低下して電位によって細胞膜上にあるナトリウム伝達通路(電位開閉型)が開口する。開口しているのはミリ秒の短時間であるが,多量のナトリウムイオンが細胞内に移行する。隣接する領域の細胞膜のナトリウム伝達通路が開口していき,脱分極の波が細胞全体に広がる。これを活動電位と呼ぶ(ニューロンでは, 神経インパルス とも呼ばれる)。
不応期
最初の刺激から0.001秒以内にニューロン(または筋繊維)に与えられた2番目の刺激は別のインパルスを誘起できない。膜は脱分極し( 上図の B ),このニューロンは不応期にある。 -70 mv の極性が再度確立されないと( 上図 C ),ニューロンは再び反応できない。
再極性化はまず,カリウム・イオンが細胞外へ促進拡散することで確立される。ニューロンが休止して,ナトリウム・イオンが細胞外から戻ってくる。
ヒトのニューロンでは,不応期は 0.001-0.002 秒である。つまり,ニューロンは毎秒 500-1000 インパルスを発することができることを意味する。
活動電位は全か無かで起こる – 悉無律 ( しつむりつ ) に従う
活動電位の強さは細胞の特性で決まっている。細胞の閾値に長く達していても,強い活動電位を起こすわけではない。ただし。強う刺激は活動電位の頻度として現わされる。
有髄ニューロン
多くのニューロンの軸索は ミエリン鞘と呼ばれる脂質の髄鞘に包まれている。髄鞘の実体は神経膠の細胞膜であって,軸索のまわりをなん層にもなって取り巻いていることが多い。 末梢神経系ではシュワン細胞という。シュワン細胞の鞘の間の部分では軸索は髄鞘をもたない。ナトリウム伝達経路( 電位開閉型 )はこの部分に限定されて認められる ( ランヴィエ結節 nodes of Ranvier と呼ばれる )。

1つの結節にナトリウム・イオンが殺到して,次のインパルスの閾値に達するに十分な脱分極が形成される。この方法で,活動電位が1つの結節から次の結節に飛び移る。これにより,非常に速い神経インパルスの伝達が可能となる。
多発性硬化症 Multiple sclerosis
自己免疫疾患 によって,ミエリン鞘が徐々に破壊される場合がある。これにもかかわらず,神経インパルスは,細胞がかつてミエリンによって保護されていた膜の一部のナトリウム伝達経路( 電位開閉型 )にイオンを追加する間伝達し続ける。
過分極
細胞は細胞膜を境に内側はマイナスに分極している。この分極の減少を脱分極,増加を過分極という。 ある神経伝達物質は神経インパルスの伝達を抑制する。 その抑制は,細胞膜内の,以下のチャンネルを開口することによって達成されている:,
- クロライド・チャネル(塩素チャネル) あるいは
- カリウム・チャンネル
それぞれの場合,チャンネルが開口して膜電位が増加する。その機序は以下の通り:
- 負に荷電した塩素イオン (Cl–) が流入し,
- 正に荷電したカリウム・イオン (K+) が流出する。
この過分極は 抑制シナプス後電位 ( IPSP ) と呼ばれている。
細胞の閾値電位が変わらないが,さらに強い刺激が必要になることがある。
例: γアミノ酪酸 Gamma amino butyric acid (GABA). この神経伝達物質は脳で見出され,以下の 2 つの機構によって神経伝達を抑制する:
- GABAA 受容体に結合すると,ニューロンの塩素チャンネルが開く。
- GABAB 受容体に結合すると,ニューロンのカリウム・チャンネルが開く
シグナルの統合
伝達物質はシナプス間隙を拡散して,次のニューロンの細胞膜の特殊な部位 ( 伝達物質を認識する特殊な膜構造,すなわち受容体のある部位 ) に働いて,細胞膜のイオン透過性を変化させる。これにより,次のニューロンのシナプス部位の膜,すなわちシナプス後膜の脱分極 ( 細胞膜の静止電位が小さくなる ) が起こると次のニューロンの興奮性が高められ,過分極 ( 細胞膜の静止電位がさらに高くなる ) が起こると興奮性が抑制される。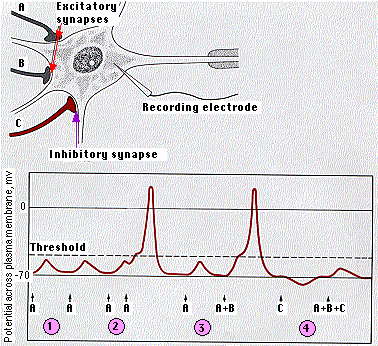
中枢神経系のニューロンは ( ニューロンのカラー写真 参照 ), とくにシナプスが多い。これらのあるものは興奮性の( 脱分極 )神経伝達物質を放出し,他のものは抑制性の( 過分極 )神経伝達物質を放出する。
受信した細胞はこれらの信号を統合する。右図は,これが運動ニューロンでどのように働くかを示す。
- 1 回の興奮性シナプスによって作成された興奮性シナプス後電位(EPSP)はニューロンの閾値に達するのに不十分である。
- しかし,連続して起こった興奮性シナプス後電位は( EPSP )は合計されて,閾値に達すると活動電位が発生する。
- 数回に亘る興奮性シナプス ( A + B ) によって作成された興奮性シナプス後電位も合計される。
- 抑制パルスの作動によって ( C ) ,ニューロンの静止電位をさらに負電荷を高める。その結果生じた抑制シナプス後電位( IPSP )も興奮性シナプス後電位( EPSP )が活動電位を引き起こす状況を抑止する。
通常,閾値に達するために必要な興奮性シナプス後電位( EPSP )の回数はここで示しているものより多い。
他の領域で抑制シグナルがあるに場合には,細胞膜の一部での脱分極で活動電位の生成が回避される。これには, 軸索小丘 axon hillock のニューロンが作用している。軸索小丘は細胞体から軸索が現れる領域である。 軸索小丘の細胞膜の部分には,
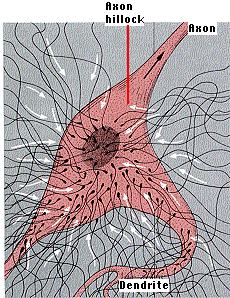
- それ自身のシナプスはなく,
- 細胞の他の領域に比べ,閾値が低い。
活動電位が軸索小丘で通常起こる。それ自身には興奮性シナプスも,抑制シナプスもないので,樹状突起と細胞体で生成された興奮性シナプス後電位( EPSP ) と 抑制シナプス後電位( IPSP )の状態が評価できる。
短時間ながら,脱分極シグナルの合計から過分極シグナルの合計を差し引いたものが,軸索小丘の閾値を超えた場合に,活動電位が起こるだろう。
ニューロンが正と負のシグナルの混じり合ったシグナルを評価する回路は急速に起こる。しかし,ニューロンが正と負のシグナルの混じり合ったシグナルを統合して判断できる長期回路をもっていることが分かっている。この長期反応にっは遺伝子活性の変化が含まれており,これにより細胞のシナプスの数や活性が変化する。
| 最初に戻る |
| メニューのページへ戻る |
February 06, 2020