| このページの内容 |
筋肉
動物は筋肉を利用して, ATP の化学エネルギーを機械仕事に変換している。脊椎動物には 3 種類の筋肉がある。
- 心筋 – 心臓を構築している筋肉。生涯を通じて,1 分間に 70 回程度収縮し,毎分約 5 リットルの血液を送り出す。
- 平滑筋 体のすべての中空器官 ( 心臓を除く内臓 ) を構築している。その収縮によって器官の大きさが縮小する。このようにして,平滑筋は:
- 動脈中の血流を調節している。
- 摂取した食物を 消化管 に沿って移動する。
- 膀胱から尿を排泄する。
- 子宮内の赤ちゃんを娩出する。
- 肺内を循環する空気の流量を調節する。
平滑筋の調節は不随意運動による。
- 骨格筋, 名前が示すように,骨格に付着する筋肉である。横紋筋 striated muscle とも呼ばれる。この筋肉は随意運動を行う。
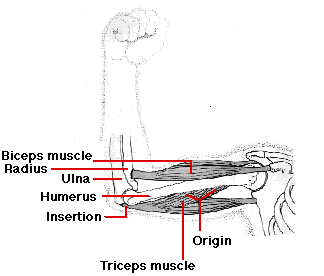
骨格筋の構造
三頭筋 ( 一端が 3 つに分岐している ) のような単独の骨格筋は,
- 骨の広い部分の “起始” に付着している。この場合は 上腕骨 humerus である。
- その他端, “停止” は次第に細くなり白色の 腱 tendon に付着する。この例の場合には 尺骨 ulna ( 前腕の骨の 1 つ ) に付着する。
三頭筋が収縮すると,”停止” は “起始” の方へ引っ張られ,腕が真っ直ぐになる。このように三頭筋は 伸筋 extensor として働く。骨格筋は収縮する場合にのみ力を発揮するので,関節を曲げるためにはもう 1 つの筋肉,屈筋 flexor が必要となる。この場合は二頭筋が前腕の屈筋である。このように二頭筋と三頭筋は拮抗筋として共に働く。 このような関節を挟んで拮抗的に働く筋肉によって 骨格 のほとんどすべての運動が可能となっている。
{kind=link}
筋繊維
1 つの骨格筋は多くの円筒状筋細胞 ( 筋繊維 という ) が束ねられたもので,個々の筋繊維は,原則として起始から停止まで伸び,途中で終わることはない。筋繊維は,多くの血管や神経が走行している結合組織によって囲まれている。
筋繊維の数は原則として生後は分裂によって増えることはない。すなわち, 1 つの筋肉に含まれている筋繊維の数は,一生を通じてほぼ一定している。筋力や筋肉量の増加は,個々の筋繊維の太さの増加ならびに結合組織量の増加によって起こる。

骨格筋を顕微鏡で側面を観察すると,その名 ( 横紋筋 ) のとおり細胞体のなかに明暗の横じまが見える。これは細胞質のなかにある収縮性フィラメントの配列によるものである。
各筋繊維には,以下のものが含まれる:
- 筋原線維 myofibril が縦方向に配列し,筋繊維の全長に亘って走行している。
- ミトコンドリア
- 発達した 小胞体
- 多数の核
多数の核があるのは,各筋繊維が多くの細胞 ( 筋芽細胞 myoblast という ) が融合して発達してきたためである。筋繊維が 1 つの細胞でないので,以下のように特別の用語が用いられる:
- 細胞膜を 筋細胞膜 sarcolemma
- 小胞体を 筋小胞体 sarcoplasmic reticulum
- ミトコンドリアを 筋粒体 sarcosome
- 細胞質を 筋形質 sarcoplasm
ただし,他の細胞で認められるものと基本的には同じ構造と機能をもっている。
- 核とミトコンドリアは細胞膜直下に位置する。
- 小胞体は 筋原繊維 myofibril 間に広がっている。
- 暗調の A 帯 と
- 明調の I 帯 からなり,
- A 帯の中央にやや明るい H 帯 があり,
- I 帯の中心を Z 帯 とよばれる線が走る。
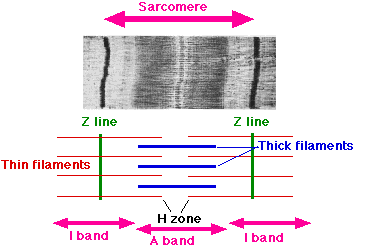 各 筋原繊維 myofibril は多くの並列した 繊維 から構成される。
各 筋原繊維 myofibril は多くの並列した 繊維 から構成される。
- 太いフィラメント thick filament は直径が約 15 nm である。この部分は,タンパク質 ミオシン myosin から構成される。
- 細いフィラメント thin filament は直径が約 5 nm である。この部分は主にタンパク質 アクチン actin からなる。ここには少量の 2 種類のタンパク質:
- トロポニン troponin と
- トロポミオシン tropomyosin がある。
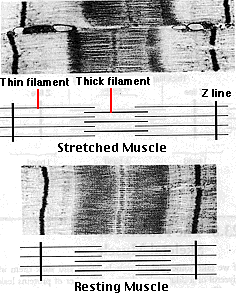
サルコメア ( 筋節 ) の構造
- 太いフィラメント は暗調の A 帯 であり,
- 細いフィラメント は Z 帯から両側に広がっている。 太いフィラメントと重なっていない部分では,明調の I 帯 となる。
- H 帯 は太いフィラメントと細いフィラメントが重なっていない A 帯の部分である。
Z 帯間の太いフィラメントと細いフィラメントの配列を,筋繊維の中の一つの単位と考え サルコメア ( 筋節 ) sarcomere と呼ぶ。筋原繊維 myofibril のサルコメア ( 筋節 ) の収縮は筋繊維が短縮することとなる。 [ この電子顕微鏡写真は 1 つのサルコメア ( 筋節 ) を示す。]
骨格筋の運動

骨格筋の収縮は神経系によって制御される。『死にかけている雌ライオン,The Dying Lioness 』 ( 約紀元前 650 年と推定されるアッシリアの彫刻,大英博物館所蔵 ) にこれが鮮明に描かれている。脊髄への損傷が,槍を受けていない後肢を麻痺させている。
この点で,骨格筋は平滑筋と心筋と異なる。筋肉はすべて神経の支配を受けている。このうち骨格筋は 脳脊髄神経 の支配を受け,平滑筋と心筋は 自律神経 ( 交感神経と副交感神経 ) の支配を受ける。
神経筋接合部
骨格筋の収縮は,脊髄内の運動ニューロンからのびた運動神経によって調節されている。運動神経繊維の インパルス が 神経筋接合部 ( シナプス の 1 種) に達すると,神経末端から伝達物質 ( アセチルコリン ) が分泌され,神経筋接合部の筋繊維の膜に作用して活動電位を発生させる。活動電位は,速やかに筋繊維の全長にわたって伝播し,筋繊維全体の収縮をひきおこす。
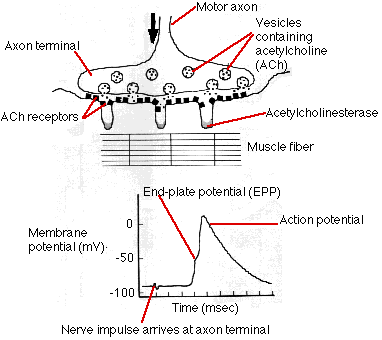
活動電位が軸索末端に達すると,多数の小胞が神経繊維上のシナプス後膜にアセチルコリンを放出する。この領域には膜貫通型の分子通路があり,アセチルコリンによってこれが開口し,ナトリウム・イオン (Na+) が流入する。
休止筋繊維内には,静止電位 は約 -95 mV である。ナトリウム・イオンの流入によって,荷電が低下し終板電位 end plate potential をつくる。終板電位が 閾値 ボルト ( 約 -50 mV ) に達すると,ナトリウム・イオンが急速に流入して,神経繊維内に 活動電位 をつくり出す。この活動電位が神経繊維全体に伝わる。
活動電位の直後は筋繊維に目に見えた変化が起こらない。この期間を潜伏期間といい,3 – 10 秒続く。
この期間に,
- 酵素 アセチルコリンエステラーゼ acetylcholinesterase が,
- 神経筋接合部のアセチルコリンを分解する ( 1 秒当たり 25,000 分子の分解速度 ) 。
- ナトリウム通路が閉鎖する。
- 新たな神経インパルスの到着のために,すべての物質が除去される。
- カリウム・イオン の流出によって,神経繊維の休止電位が回復する。
休止電位が回復するのに極めて短い期間が必要である ( 1 – 2 msec) 。これを 不応期 refractory period と呼んでいる。
強縮
収縮の過程は約 50 msec かかり,筋繊維の弛緩にはさらに 50-100 msec かかる。収縮と弛緩に必要な時間よりも不応期が短いので,頻繁に刺激を受けた時には ( たとえば,1 秒間に 50 回くらいの刺激 ) 筋繊維は収縮したままの状態を維持できる。そのような持続性の収縮を 強縮 tetanus を呼んでいる。
 右図では,
右図では,
- 1 秒間に 1 回刺激を与えると,筋肉は単収縮 ( 単一の活動電位によっておこる短い収縮 ) を起こす
- 1 秒間に 5 回ならびに 10 回刺激を与えると,個々の単収縮が融合し始める。この現象を クローヌス ( 間代性痙攣 ) clonus という。
- 1 秒間に 50 回刺激を与えると,筋肉は持続性の強縮を起こす。
収縮と弛緩の周期性を完了するの要する時間より不応期が非常に短いので,クローヌスと強縮が起こすことが出来る。単収縮で見られるよりもクローヌスと強縮で収縮量が大きいことに注意して欲しい。
われわれの身体の運動はほとんどすべてが骨格筋の強縮によるものである。
滑り説
,太いフィラメントはミオシンからなる。アクチンフィラメントは球状のアクチン分子が二重ラセンをなして重合したものである。ミオシン分子はふくらんだ 2 個の頭部と棒状の尾部をもつ。尾部は束になって太いフィラメントの軸を形成し,頭部は側方に突き出て側枝となっている。筋収縮の際 2 種の筋フィラメントは能動的に筋繊維の長さを短くする方向にたがいに滑り込んでいく。 太いフィラメントのミオシン分子はふくらんだ 2 個の頭部と棒状の尾部をもつ。 ミオシン頭部 が,以下に結合する:
- 細いフィラメント の アクチン 分子 と
- ATP
筋繊維の活性化により,ミオシン頭部がアクチンに結合する。アロステリック 変化が起こって,細いフィラメントをわずかに ( ~10 nm ) 引き寄せ,太いフィラメントのそばを通り過ぎる変化が生じる。それから結合が解離して ( これには ATP が必要 ),細いフィラメントは元の位置に復帰し,これを周期的に繰り返す。各フィラメントは滑り合うのであって,それぞれが短くなったり太くなったり,あるいは折り畳まれているのではない。
電子顕微鏡を用いた研究から,このモデルが提唱されている。
筋肉が収縮する時,
- Z 帯がお互いに近接する。
- I 帯の幅が減少する。
- H 帯の幅が減少するが,
- A 帯の幅には変化が見られない。
逆に,伸張する時には,
- I 帯と H 帯の幅が増加するが,
- A 帯の幅には変化が見られない。
以上のことが,電子顕微鏡写真で観察できる。これが,筋肉の収縮はアクチンとミオシンの間に滑込みが起こるという,いわゆる “滑り説” である。
筋収縮に対するカルシウム・イオンの役割
筋収縮は,筋繊維のまわりの微量のカルシウム・イオン ( Ca2+ ) の増減によって調節される。
- 休止状態の筋肉繊維では, Ca2+ が筋小胞体に貯蔵されている。
- 筋繊維の細胞膜は多くの部位で管状に内部におちくぼみをつくっており,これを 横行小管系 ( T 系 ) transverse system という。
- 横行小管は筋小胞体と多くの部位で密接しているので,
- 筋繊維細胞膜におこる活動電位あるいは脱分極の影響は筋小胞体にただちに伝えられ,
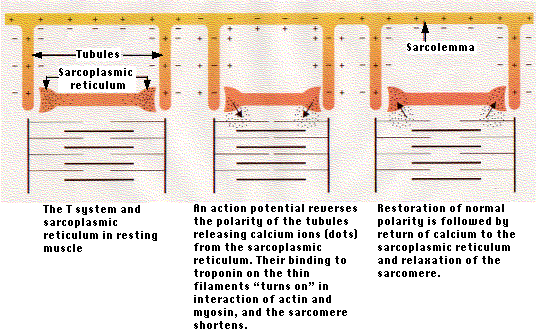
- その結果,筋小胞体内腔の Ca2+ が筋細胞質中に放出される。
- Ca2+ が太いフィラメントと細いフィラメントの周囲に拡散して,そこで
- 細いフィラメント上の トロポニン troponin と結合する
- これが,アクチンとミオシンの相互作用のスイッチを入れ,サルコメア(筋節)が収縮する。
- 活動電位の速度のために ( ミリ秒 ),活動電位がすべての横行小管系の終末にほとんど同時に到達するので,すべてのサルコメア ( 筋節 ) が一斉に収縮する。
- この過程が完了すると,が Ca2+ ATPase が使われ,カルシウム筋小胞体に回収される。
等張力性収縮と等尺性収縮
筋肉の収縮中,筋肉に加わっている力 ( あるいは荷重 ) が一定に保たれている場合の筋肉の収縮を 等張力性収縮 という。
一方,筋肉の両端を固定して短縮できないようにして収縮させたとき,このような収縮を 等尺性収縮 ( 長さが一定 ) という。
運動単位
骨格筋は,自発性に収縮することはなく,運動神経を通って適切な頻度で送られてくるインパルスによって収縮する。
運動神経繊維を出している神経細胞は脊髄の灰白質の前角にあり,a 運動ニューロンという。運動単位 とは反射および随意収縮の生理学的単位であり,1 個の a 運動ニューロンとその支配下にある筋繊維を一緒にしたものを指す。一般に 1 つの筋肉に含まれる筋繊維の数は,その筋肉を支配する a 運動ニューロンの数よりずっと多い
運動単位の大きさは支配する神経繊維と筋繊維の数の比,つまり神経支配比で示される。
- 正確で細かい急速運動をする眼筋などは 1 対 3 – 8 で神経支配比は小さい。
- 咽頭を支配する筋肉の運動単位も 1 対 2 – 3 と小さい。
- これに対して,大きな収縮力を発生する下腿の筋肉では神経支配比は 1 対 150 程度と大きい。
- 子牛の腓腹筋のような筋では,1000 – 2000 と大きい ( 筋肉に均一に分散している ) 。
筋収縮のエネルギー
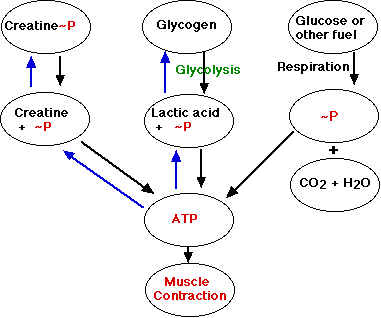 ATP が筋収縮の直接のエネルギー源である。筋繊維は十分な ATP があれば数回の単収縮できるが,ATP の貯蔵が必要となる。 ATP 貯蔵のための高エネルギーリン酸の 3 つの供給源がある。
ATP が筋収縮の直接のエネルギー源である。筋繊維は十分な ATP があれば数回の単収縮できるが,ATP の貯蔵が必要となる。 ATP 貯蔵のための高エネルギーリン酸の 3 つの供給源がある。
クレアチンリン酸
クレアチンリン酸内の リン酸基 が,ATP のような高エネルギー結合として付加される。クレアチンリン酸は ATP から高エネルギーリン酸を誘導し,それを ADP に戻し ATP を形成する。
{kind=link}
クレアチンリン酸 + ADP <–> クレアチン + ATP
筋繊維内のクレアチンリン酸の貯蔵は,ATP のそれに比べ10倍以上も多い。
a name=”glycolysis”>
<解糖
骨格筋は約 1% のグリコーゲンを含有する。 筋繊維は解糖によってこのグリコーゲン分解することができる。 解糖 によって,生産された乳酸分子あたり 2 モルの ATP を生産する。多くはないが,呼吸によって十分な酸素を受け取らない場合に,筋肉の機能を保つのに十分である。しかし制限があるので,最終的に筋肉は細胞呼吸に依存する。
細胞呼吸
細胞呼吸 は長い活動に従事するための ATP 要求を満たすためばかりでなく(このため呼吸も深く速くなる),前に生産された乳酸からグリコーゲンの再合成するためにも必要である(運動停止後しばらく深呼吸が続く)。体はこの酸素消費に対して生理的に反応することになる。
2 種類の筋繊維
筋繊維には白筋繊維と赤筋繊維の 2 種類がある。前者は,太くて,ミトコンドリアが少なく,敏速な収縮を起こすが疲れやすい。後者は,細くて,で,,。
タイプ I 筋繊維(赤筋)
- ミトコンドリアが豊富,
- ATP の生産は 細胞呼吸 による ( 酸化系酵素に富む )
- おそいが持続的な収縮を行う
- ミオグロビンが豊富,したがって赤筋
- 細く,遅延型の運動ニューロンが支配する
タイプ II 筋繊維 ( 白筋 )
- ミトコンドリアが少なく,
- グリコーゲンが豊富
- ATP の生産は 解糖 による
- 解糖に依存しているため疲れやすい
- ミオグロビンが少ないので色が白っぽい
- 太く,敏速な運動ニューロンが支配する
- 敏速な収縮を起こす
ほとんどの骨格筋は両者の筋繊維を混合してもつが, 運動単位 は 1 つでどちらかが含まれる。両方があることはない。
両タイプの筋肉の比率を,筋肉トレーニングによって変えることが出来る ( タイプ I 筋繊維を多く製造する ) 。
心筋
心筋はある意味で骨格筋に似ている。横紋筋であり,各細胞はアクチンとミオシンの滑り繊維からなるサルコメア ( 筋節 ) を含む。
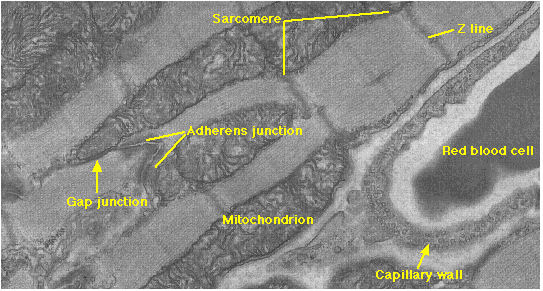
しかし,心筋は多くの独特な特徴を持ち,血液を送り出している。
- 単核の分枝のある心筋繊維 myofibril ( 細胞 ) が網目状につながったもので,
- 心筋細胞の境界には 接着結合 という細胞の接着装置が発達していて,心筋細胞どうしが収縮のときに離れないようになっている。
| 上の電子顕微鏡写真では,接着結合 adherens junction ならびにこのページで紹介したいろいろな情報が観察できる。 |
- 心拍を生み出す活動電位は心臓自体で生み出される。運動神経 ( 自律神経系 ) が心臓に達しているが,その効果は,心臓本来の心拍の早さと強さを調整する。神経が破壊されても,心臓は心拍を続ける。
- 心筋細胞の間には ギャップ結合 という結合装置があり,これによって興奮が伝えられる。ギャップ結合は小分子物質やイオンを通過させて細胞間の情報伝達にあずかる装置である。
- 心筋の 不応期 は収縮期と拡張期にかかる時間よりも長い。したがって,強縮は起こらない。
- 心筋は骨格筋よりもミトコンドリアの供給を豊富に受ける。これは ATP を細胞呼吸により依存していることを示す。
- 心筋はほとんどグリコーゲンをもたない。酸素供給が限定されてしまうので,解糖にほとんど依存していない。
- 酸素の豊富な血流が停止すると心臓は極めて急速に損傷を受ける。つまり,心臓発作である。
平滑筋
平滑筋細胞は 1 個の紡錘形細胞から成る ( 長さは 20 – 200 mm,太さは約 5 mm で,細胞の中央部に 1 個の核をもつ ) 。骨格筋や心筋などと異なり,横紋をもたないことから,この名がある。
ミオフィラメントとしてはアクチンやミオシンが存在するが,横紋筋に比べて量も少なく,規則的配列も示さない。 太いフィラメント(ミオシン)と細いフィラメント ( アクチン ) は細胞膜付近に固定されている( 中間径フィラメント によって ) 。
平滑筋は ( 心筋と同様に ),その刺激を運動ニューロンに依存していない。しかし,自律神経系の運動ニューロンが平滑筋に達し,これが分泌する神経伝達物質 ( たとえば,ノルアドレナリンや一酸化窒素 ) に依存して,平滑筋を刺激,または弛緩させている。
平滑筋は,以下の刺激によっても収縮する:
- パラクリン ( 傍分泌 ) 刺激によって放出される他の物質によって,
- 例: ヒスタミン の放出 – 気管の平滑筋の収縮を起こす ( 喘息の発作など )
- 血中のホルモンによって,
- 例: オキシトシン は子宮筋の収縮を起こし,胎児の娩出などに重要
平滑筋の収縮は骨格筋のそれに比べゆっくりとした傾向がある。これも緊張と呼ばれるが,その機構は骨格筋と異なるようである。
筋肉の病気
a name=”MD”>
<筋ジストロフィー ( MD )
ミオシン,アクチン,トロポミオシンおよびトロポニンはともに,筋繊維の蛋白質の 3/4 以上を占める。その他 20 種ほどのタンパク質が含まれる。これらのタンパク質はサルコメア( 筋節 )中の筋繊維の組織化に関わり,また細胞膜および細胞外基質へ筋節を結合させるのに働いている。これらのタンパク質をコード化する遺伝子の突然変異によって,筋肉中に正常に機能しないタンパク質やいろいろな欠陥を生じることがある。
筋ジストロフィー症の一番多い原因は ジストロフィン dystrophin 遺伝子の突然変異によるものである。 .
ジストロフィン dystrophin 遺伝子は巨大で,230 万塩基対以上の DNA 分子に広がった 79 個のエクソンを含む。これはヒトゲノム ( 3 x 109 bp ) の約 0.1% に相当し, 大腸菌のゲノムサイズ のほぼ半分である。
- デュシェンヌ型筋ジストロフィー Duchenne muscular dystrophy ( DMD )遺伝子が巨大なために,部分的な 欠失 が疑われる。リーディング・フレームが変化し,機能的なジストロフィンが合成されない。
- ベッカー型筋ジストロフィー Becker muscular dystrophy ( BMD )欠失のために,特定のエクソンが取り除かれ,短縮したタンパク質が生成される。
ジストロフィン遺伝子は X 染色体上にある ( X-連鎖 形質)。したがって,これらの 2 つの遺伝病は典型的な伴性遺伝に従う。
重症筋無力症
重症筋無力症は 神経筋接合部 に影響する自己免疫疾患である。患者は標準より小さな 終板電位 ( EPP ) を持っている。繰り返された刺激によって EPP があまりにも小さくなるので,活動電位を引き起こすことがでず,筋繊維が収縮を中止してしまう。アセチルコリンエステラーゼの抑制剤の投与によって,一時的に収縮を回復できる ( より多くのアセチルコリンが接合部に残るため ) 。
重症筋無力症を持った患者は,通常の 神経筋接合部 で見つかったアセチルコリン受容体の数が 20% 程度しか持っていない。受容体の消失は,受容体に対する 抗体 によって引き起こされたようである。そのいくつかの証拠:
- アセチルコリン受容体に対する抗体で免疫することにより,実験の動物に重症筋無力症に似ている疾病を引き起こすことが可能である。
- 抗-アセチルコリン受容体抗体が,患者の血清中に検出される。
- 患者の血清を実験動物に投与すると,重症筋無力症の症状を示す。
- 重症筋無力症の母親の新生児は出生後まもなくしてこの病気の症状を示す。これは,妊娠時に胎盤経由で母親の抗体が移行した結果と考えられる。
アセチルコリン受容体に対する自己免疫抗体をつくってしまう理由は分かっていない。
心筋症
心臓の筋肉は骨格筋のように,アクチンとミオシンに加えて多くのタンパク質を含んでいる。これらの遺伝子の突然変異は,心臓の壁を弱めるような原因となる。そのため,やがて拡張してしまう。これらの疾病に含まれる遺伝子は,次のものをコード化している:
- アクチン
- 2 種類のミオシン
- トロポニン
- トロポミオシンtropomyosin
- ミオシン結合タンパク質 C
病気の程度はそれを起こす遺伝子の突然変異 ( これまで100以上が同定されている ) によって様々である。 ある突然変異のために,健康で活動的な成人が突然死に至る心疾患を発症する危険性は十分にある。
| 最初に戻る |
| メニューのページへ戻る |
February 04, 2020