ポリジーンと多因子性遺伝
| このページの内容 |
複数の遺伝子によって制御されている形質
メンデルのエンドウ豆の高さは 1 組の遺伝子によって決まるが,ヒトの身長を決めるのには多くの遺伝子が関与している。 2 組以上の遺伝子によって決定される形質は ポリジーン形質 polygenic trait と呼ばれる。
- エンドウ豆の「 高い 」ものと「 低い 」ものは, 2 つの明確な表現型に区別され,これらを 不連続変異 という。
- ヒトの身長を,いくつかの表現型に分けるのは不可能で,これは 連続変異 の代表的な例である。
集団内のヒトの身長は重複した値をとり,しかも連続した表現型( 値 )を示す。段階的な表現型を示す形質は,通常複数の遺伝子によって支配される。
| 連続形質の別な例 |
不連続形質と連続形質の違いを理解することは,遺伝学を学ぶ上で重要である。遺伝子同士が相互作用を及ぼし,また環境とも相互作用をもつという概念に基づいている。
複数の遺伝子によって支配され,また環境要因とも相互作用を示す形質を 多因子性形質 multifactorial trait と呼ぶ。 このページでは,複数の 遺伝子座 loci によって支配されている形質について学ぶ。さらに,環境要因 が関与する形質についても紹介する。
多因子性遺伝の場合には,遺伝子の影響の程度は 遺伝率 を算出することによって評価する。この概念については,一卵性双子の例を用いて学ぶ。
このページの最後では,ヒトにおけるいろいろなポリジーン形質についてふれる。
ポリジーンと表現型の変異
ポリジーンによる遺伝様式では,表現型の連続的な分布はいくつかの明確な性質をもつ。
- 数値で定量化される形質である ( 身長や体重など ) 。
- 複数の遺伝子が関与するが,各遺伝子は相加的に働く。しかもこの対立遺伝子の影響は小さい。
- ポリジーン遺伝の表現型発現 ( 変異 ) は多種多様である。この変異は個体レベルではなく,集団レベルで解析される。
ポリジーンによる遺伝様式は,ヒトの遺伝学で非常に重要な概念である。
- 身長
- 体重
- 皮膚の色
- 知能
などの形質はポリジーン支配下にある。
 さらに,
さらに,
- 先天性奇形
- 遺伝病 ( 糖尿病,高血圧,行動障害など )
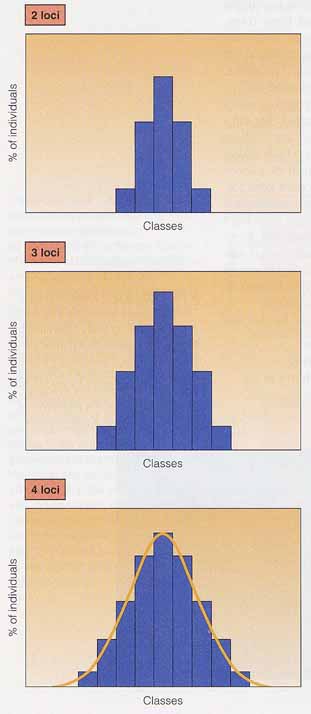
もポリジーン形質または多因子性形質である。 表現型と遺伝子を 2 個,3 個,ならびに 4 個含む形質の雑種第 2 代 ( F2 ) 比の分布を左図に示す。2 種類の遺伝子が 1 つの形質の発現に関与していれば,F2 では表現型 5 つの階級に分けられる。すなわち,優性の対立遺伝子を 4 個,3 個,2 個,1 個,0 個をもつ階級である。F2 比,1:4:6:4:1 は各表現型に関わる遺伝子型の組み合わせによって決まる。4 個の優性対立遺伝子をもつホモ接合体 ( AABB ) が最大の組み合わせであり,0 個の優性対立遺伝子をもつホモ接合体 ( aabb ) が最小の組み合わせである。
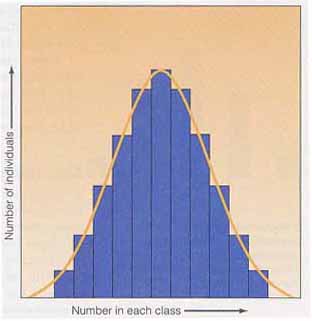
最も度数の多い ( 6/16 ) 階級は,2 個の優性対立遺伝子をもつ遺伝子型の組み合わせである ( 1AaBb,4AAbb,1aaBB ) 。形質に関与する遺伝子の数が増えるにしたがい,表現型の変動幅は増大する。また,環境要因の関与により,表現型が融合して,連続分布を生み出すことになる。 形質に関与する遺伝子座の数が増えるにしたがい,表現型の階級の数が増加する。階級の数が増加すれば,各階級間の表現型の差は小さくなる。このことは,環境要因による影響が各階級間の小さな差を乗り越えてしまう機会が多くなることを意味する。たとえば,日光に当たると皮膚の色が変化し,表現型の差が不明瞭になってしまうことがある。ポリジーン遺伝の表現型値は度数分布図で良く表わされる( 右図 ) 。
1. ポリジーン遺伝の相加的モデル
 ポリジーンが形質発現にどのように関与しているのか,遺伝子型が表現型の変異にどのように関与しているのかを説明するための,ポリジーン遺伝のモデルを次の条件で考えてみよう。
ポリジーンが形質発現にどのように関与しているのか,遺伝子型が表現型の変異にどのように関与しているのかを説明するための,ポリジーン遺伝のモデルを次の条件で考えてみよう。
- 複数組の遺伝子で形質が支配されているとする。ここでは 3 組の遺伝子を想定し,それぞれが 2 個の対立遺伝子をもつとする( A,a, B,b, C,c )。
- 各々の優性対立遺伝子は表現型に “均等” に関与し,劣勢遺伝子は全く関与しないこととする。
- 各々の優性対立遺伝子の表現型に対する影響は小さく,”相加的” なものとする。
- この形質を制御する遺伝子は連関せず,それぞれ独立に関与することとする。
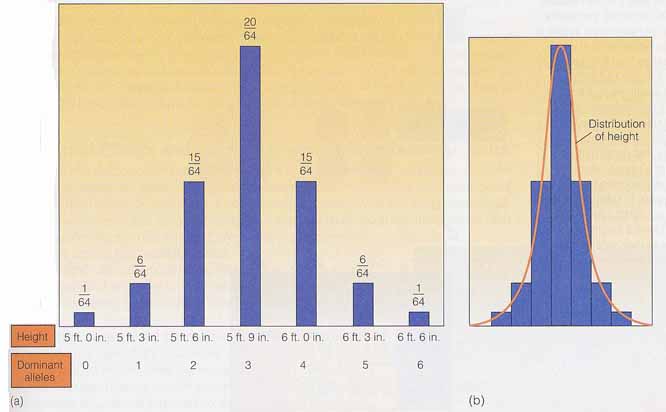
このモデルを身長の遺伝に当てはめてみよう。 ここでは,基礎値 5 フィート( 150 cm )に対し,優性遺伝子 1 個あたり 3 インチ( 7.5 cm )ずつ高くなると仮定する。劣性の対立遺伝子 a,b,c は基礎値に対して何の追加もないこととする。
- 遺伝子型 aabbcc をもつ人は身長が 150 cm であり,
- 遺伝子型 AABBCC をもつ人は 195 cm ということになる。
- 環境要因がすべての遺伝子型に均等に作用するとすれば,6 個のうち 3 個の優性遺伝子をもつ人の平均身長は 172.5 cm ということになる。
右図 ( a ) は遺伝子型頻度とそれらの表型値を示す。3 つの対立遺伝子を想定したので,7 つの表現型の階級がある。すなわち,6 個,5 個,4 個,3 個,2 個,1 個,0 個の優性遺伝子をもつ階級である。
現実には,環境要因が作用して,階級の差異を不明瞭なものにしてしまうので,実際経験するような連続的な変異と示すことになる( b )。
最も頻度の高い表型値が身長の平均値 172.5 cm ということになる。表型値の両端に行くにつれ,頻度は低下し,1/64 の人が 150 cm または 195 cm の身長を示す。
この例や一般的なポリジーン形質では,
- 遺伝子型が身長に対する 遺伝的能力 を示す。
- 遺伝子の能力が完全に発揮されるかどうかは 環境要因 による。
- 発育時期に栄養が十分でなかった場合には,遺伝的な身長にも達しないこともあるし,
- 他方十分な栄養の元で発育した場合には,遺伝的な身長以上の背の高さになることもある。
2. 表現値の平均化
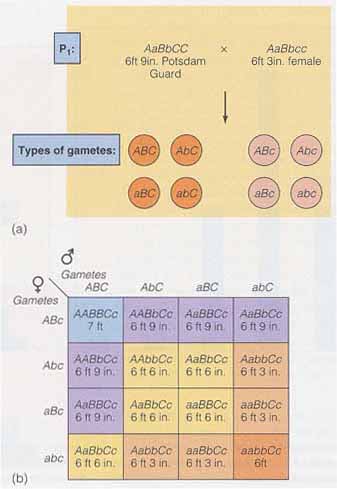
ポリジーン形質の特徴の一つは,最大値( または最小値 )をもつ個体を親とした場合,ほとんどの子供は極端な値をとらず,表型値は平均化することである。この現象は平均値の逆行 regression of the mean と呼ばれる。これに関する例を,下の両図に示す。 平均値の逆行は,遺伝子の相互作用や環境効果によって生じる遺伝子の 優性効果 と 相加的効果 が大きいと,顕著に認められる。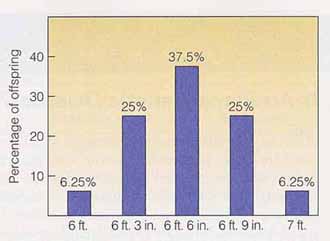
ポリジーンと環境:多因子性形質
個々人の表型値は,どれだけが遺伝によるもので,どれだけが環境によるものなのだろうか?
各個人は独特の遺伝子型をもち,独特の環境条件の組み合わせのもとにおかれているので,表現型の遺伝的な要素と環境的な要素を量的に評価するのは不可能である。しかし,ちょっと見方を変えて考えて見よう。
1. 閾値効果と多因子性形質の発現
遺伝子型と環境の相互作用を推定するのは難しいが,家系調査によってそれらの相互作用が起こることが知られている。ある多因子性形質は表現型の連続的な分布を示さない。すなわち,影響を受けるか受けないかのどちらかである。先天性の奇形がその例である。
遺伝病を引き起こす “不都合の程度” は,集団で見ると正規分布を示す。この “不都合の程度” は遺伝子型で表わすことができるが,特定の数の遺伝子型においてのみ,表現型が発現する。
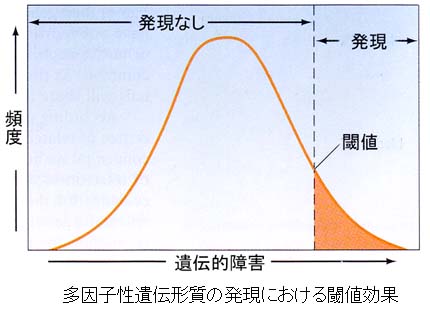 右図に示したように,疾病素因は遺伝子の数に依存し,それぞれの遺伝子が相加的に障害発生に関与する。特定の遺伝的な閾値を超えた “不都合の程度” を持つ個人が,特定の環境条件におかれた場合に遺伝病になる例である。言い換えると,環境条件が遺伝的な疾病素因をもつ人に最も大きな影響をもたらす例である。
右図に示したように,疾病素因は遺伝子の数に依存し,それぞれの遺伝子が相加的に障害発生に関与する。特定の遺伝的な閾値を超えた “不都合の程度” を持つ個人が,特定の環境条件におかれた場合に遺伝病になる例である。言い換えると,環境条件が遺伝的な疾病素因をもつ人に最も大きな影響をもたらす例である。
2. 遺伝子型と環境の相互作用の推定
では,どのようにして遺伝子型と環境の相互作用が測定できるのだろうか。そのためには,集団の構成員についてみるのではなく,集団における表現型全体の変異を検討する必要がある。
表現型の変異は次の 2 つの要因に分けて考えることができる。
- 集団構成員の異なる 遺伝子型の影響
- すべての遺伝子型が発現する際の異なる 環境要因の影響
表現型の変異をもたらすこれらの要因を評価するために,遺伝率 heritability として知られている概念が用いられる。
遺伝率は表現型の変動に対する遺伝的貢献度を表わす
- 異なる遺伝子型によって生ずる表現型の変異は 遺伝的変異 genetic variance という。
- もし同一の遺伝子型をもつ個体間で表現型の変動がみられれば,この変異は 環境変異 environmental variance という。
形質の遺伝率は,記号 h2 で表わされ,表現型全変異に対する遺伝的な差異によって生じる変異の割合である。
特定の形質に対して絶対的な遺伝率が存在することはなく,遺伝率は変動する。どの集団を問題とするのか,あるいは測定時に関与する環境変異などを含むいくつかの要因によって,遺伝率は影響を受ける。
遺伝率は集団に認められる現象で,血縁関係がある個体を含む集団で分析される形質に対する遺伝的な貢献度を示す値である。
一般的に,遺伝率が
- 高ければ( h2 = 1 ),表現型の変異はもっぱら遺伝的な影響によるもので,環境の影響はほとんど無い,
- 低ければ( h2 = ~0 ),観察されている表現型変異に対する遺伝的な影響はほとんど無く,環境の影響が高い,
ことを示す。
1. 遺伝率の推定値は既知の血縁関係の程度から求められる
血縁個体によって共有される遺伝子の割合を知ることができるので,遺伝率は血縁個体間での観察結果から推定できる。
たとえば,
- 両親と子供は 1/2 の遺伝子を共有する。
- 祖父母と孫では 1/4 の遺伝子を共有する。
特定の表現型値の 相関係数 correlation coefficient を求め,これらの血縁関係で補正する。
子供における特定形質の母( または父 )親のそれに対する相関係数 r ( または回帰係数 ) を求めると,
 |
h2 ≒ 2r
となる。 たとえば,遺伝率が 0.72 ということであれば,その集団でみられる表現型変異の 72% が遺伝的な影響によって生じるということを意味する。
2. 指紋の遺伝率を推定してみる
遺伝子と環境の相互作用のために,多因子性形質の遺伝率を算出することは困難である。しかし,指紋がうまく遺伝率が算出できている 1 つの例である。
指紋は胎齢 3 ヵ月( 6 – 13 週 )で決まる。指紋はいくつかの遺伝子によって制御され,この短い期間に環境によって影響を受ける。一卵性双子を含め,すべての人間が,独特の指紋をもつ。一卵性双子は同一の遺伝子セットを共有し,同一の子宮内で同時に発育するにもかかわらず,僅かに異なる微環境のために,異なる指紋を形成する。
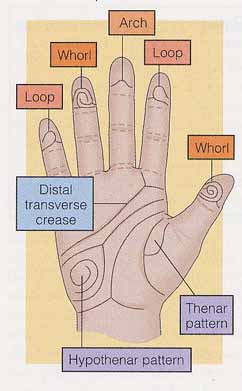
指紋は皮膚隆起線の紋様である。指紋が発達するにつれ,隆起線は明瞭なパターンに決まってくる。同じように,皮膚隆起線は手のひら,足指,足の裏にも形成される。これらのパターンの解析をすることは皮膚紋理学 dermatoglyphics と呼ばれる。手のひらの模様( 屈曲襞 flexion crease という )は手相鑑定の感情線,生命線などと呼ばれているが,同じような時期に形成される ( 右図 )。
指紋のパターンは,その形状と隆起線の数で分類される:arch 弓状紋,loop 蹄状紋,whorl 渦状紋の 3 つである( 右図 )。隆起線の数が,表現型の変異と遺伝率を研究する上で最も有用な指紋の特徴である。容易に,かつ客観的に測定できる。また,一度確定すれば,社会的または環境的要因に左右されることはない。
相関係数を利用して,男性 825 名,女性 825 名の全隆起線数 ( TRC, total ridge count ) を調査した報告がある( 表 1 )。その結果,いろいろな血縁関係にある個体間の観察値と期待値がほぼ一致していることから,TRC は概ね遺伝的支配下にあり,環境要因の働きは微々たるものであると考えられる。
両親とその子供の隆起線数の分析により,隆起線数の遺伝率は以前から h2 = 0.95 と推定されてきた。遺伝率の値が 0.95 ということは,指紋の場合の表現型変異のほとんどが遺伝的なものであり,両親から子供へ伝達されたことを示す。
| 表 1. 隆起線数と血縁個体間の相関関係 | ||||
|---|---|---|---|---|
| 血縁関係 | 検査組数 | 相関係数観察値 | 期待値 | 遺伝率 |
| 母親と子供 | 405 | 0.48 ± 0.04 | 0.50 | 0.96 |
| 父親と子供 | 405 | 0.48 ± 0.04 | 0.50 | 0.98 |
| 夫 婦 | 200 | 0.05 ± 0.07 | 0.00 | – |
| 兄弟姉妹 | 642 | 0.50 ± 0.04 | 0.50 | 1.0 |
| 一卵性双子 | 80 | 0.95 ± 0.01 | 1.00 | 0.95 |
| 二卵性双子 | 92 | 0.49 ± 0.08 | 0.50 | 0.98 |
| 出典: Quantitative genetics of fingerprint patterns, by S.B. Holt ( 1961 ). Br. Med. Bull., 17, 247-250. より改変 |
||||
すなわち,隆起線数にみられる表現型変異の 95% が遺伝子型の差異によるものであることを意味する。ほんの僅かな非遺伝的な変異が一卵性双子における指紋パターンが異なる原因と考えられる。
双子に関する研究と多因子性形質
上述のように,血縁個体における表現型値の相関係数を算出することにより,遺伝率の推定値が得られる。しかし,この方法には 1 つ大きな問題がある。
遺伝的な関係が近ければ近い程,当該の血縁個体は共通の環境を共有することが多いので,たとえば,両親と子供の表現型の類似性は遺伝子の半数セットをもっているためなのか,あるいは同様の環境に暮らしているためなのか判断で難しい。
遺伝的な影響と環境の影響を完全に分離して検討するために:
- 双子について検討する。
一卵性双子は同一の遺伝子型を共有している。一卵性双子が誕生してから,別々の環境で育てられていれば,遺伝子型は同一であるが環境因子が異なることになる。 - これとは逆に,血縁関係のない養子の形質を同一家族内の本来の子供の形質と比較する。
この場合,環境要因は一定であるが,遺伝子型が著しく異なることになる。
このように,双子に関する研究と養子に関する研究はヒトの遺伝率を算定する上で重要な手段となっている。
| 表 2. 一卵性双子と二卵性双子の一致性 | ||
|---|---|---|
| 形質 | 一致率 ( % ) | |
| 一卵性双子 | 二卵性双子 | |
| 血液型 | 100 | 66 |
| 眼の色 | 99 | 28 |
| 精神発達 | 97 | 37 |
| 髪の色 | 89 | 22 |
| ダウン症 | 89 | 7 |
| 利き腕 | 79 | 77 |
| てんかん | 72 | 15 |
| 糖尿病 | 65 | 18 |
| 結核 | 56 | 12 |
1. 双子と一致性
双子間で表現型の相違を比較する簡単な方法は,量的形質よりも悉無形質を用いることである。双子の両個体がある形質を示せば一致性をもち,一方の個体のみ保有する場合は不一致ということである。一卵性双子は通常 100% 同じ遺伝子をもつが,二卵性双子は平均 50% の同じ遺伝子をもつ。
遺伝的に決定される形質の場合,一卵性双子における相関が二卵性双子におけるものより高いはずである。もしその形質が遺伝子によって完全に制御されていれば,一致性は一卵性双子で 1.0,二卵性双子では 0.5 に近似するはずである。
この問題を検討する上で,一卵性双子と二卵性双子における一致性の差異の程度が重要である。差が大きければ大きい程,遺伝率が高いということになる。いくつかの形質に関する双子の一致性を表 2 にまとめた。
一致率は統計的な手法により,遺伝率に変換できる。この方法で算出した遺伝率を表 3 に示す。遺伝率は相対的な値であり,測定された集団とその時点で効果を及ぼしている環境要因の影響下で有効である。したがって,ある集団の遺伝率を他の集団の同じ形質の遺伝率と比較することはできない。それは, 2 つの集団が遺伝子型も環境変異も異なるからである。
2. 肥満症の研究に双子を用いる
| 表 3. 双子における肥満症の推定遺伝率 | |
|---|---|
| 条 件 | 遺伝率 |
| 小児期の肥満症 | 0.77 – 0.88 |
| 成人期の肥満症 ( 45 歳時の体重 ) | 0.64 |
| 成人期の肥満症 ( 20 歳時の 体格指数 ) | 0.80 |
| 成人期の肥満症 ( 軍隊入隊時 ) | 0.77 |
| 双子における肥満症 | |
| 男 性 | 0.70 |
| 女 性 | 0.66 |
双子や養子の研究から得られた遺伝率の推定値は多因子性形質を研究する間接的な方法である。これらの研究は原因と結果を直接示すというよりも,相関関係に基づいており,次節に示すような多くの不確定因子の影響を受ける。
肥満症は遺伝的に伝わる形質であり,アメリカ合衆国の成人の 61% までが太り気味または肥満であると推定されている。これらの人たちは,高血圧症,コレステロール高血症,冠状動脈疾患ならびに成人発症型糖尿病などの疾病を発症する恐れがある。
肥満症の遺伝率を推定するのに双子の研究が用いられた。1917 年から 1927 年に生まれ,兵役についた 1,974 組の一卵性双子と 2,097 組の二卵性双子を用いた研究がある。入隊時に測定した体重と身長が,同様に 25 年後のデータと比較された。
体格指数 body-mass index ( BMI,体重 kg を身長 m の平方根で割った数字 ) と理想の体重-身長表を基に肥満度が測定された。表 4 は,5 段階の肥満度に対してこの調査から求められた一致率を示す。
入隊時には,すべての肥満度において一卵性双子の一致率が二卵性双子のそれに比べ非常に高い。また,25 年後でも同様に高かった。表 3 はこれらの調査から求められた遺伝率である。肥満症に対する遺伝率は高く,この状態が遺伝的に強力に支配されていることが示唆される。
| 表 4. 双子における肥満症の一致率 | ||||
|---|---|---|---|---|
| 体重超過 % | 入隊時の一致率 | 25 年後の一致率 | ||
| 一卵性双子 | 二卵性双子 | 一卵性双子 | 二卵性双子 | |
| 15 | 61 | 31 | 68 | 49 |
| 20 | 57 | 27 | 60 | 40 |
| 25 | 46 | 24 | 54 | 26 |
| 30 | 51 | 19 | 47 | 16 |
| 40 | 44 | 0 | 36 | 6 |
3. 双子を用いる研究は間接的である:その問題点
他の双子を用いる研究と同様に,この研究の結果は相関関係を示すものであり,原因と結果の直接性を明らかにするものではない。結果の解釈に影響するこの種の研究の問題点を考えてみる。
- この調査には入隊時の身体検査に合格した人だけが含まれ,不合格になった極端な肥満を示す人は除外されている。
- したがって,スタート時点でこの調査の双子の集団が肥満症の軽い方向へ偏ってしまった。
- 肥満の人が除外されたため,この調査の肥満に対する遺伝率は過小評価されたしまった。
- この調査は女性,子供,あるいは他の原因で兵役から除外された男性を含んでいない。
したがって,一般的な結論を引き出せない。 - この調査はダイエットなどのような,環境要因の働きを直接評価していなかった。
しかし,これらの限界があったとしても,肥満症が遺伝の強い影響下にあることを示すに十分高い遺伝率が認められている。
他の同様の調査も,それぞれ限界があるが,十分に計画された双子を用いる研究によって得られて結論は,ヒトの多因子性形質の遺伝率の概要をとらえるのに有益な手段である。
4. 肥満症の遺伝率
遺伝子と環境の影響を分離し,またそれ以外の要因を制御する最も効果的な方法は,異なる環境条件下で育てられた一卵性双子と二卵性双子を調査することである。
このような条件で実施された肥満度に関する調査結果によると,肥満度の遺伝率は男性で 0.70,女性で 0.66 と算出されている( 表 3 )。すなわち,観察された表現型の変異( この場合,肥満症の変異 )の 66% から 70% が遺伝子型の違いによるものであり,肥満の状態が遺伝的な要因に強く関連していることが示されている。
5. 肥満症の遺伝学的な手掛かり
遺伝率の推定は表現型レベルで行われる遺伝学的な解析の一手法である。しかし,この手法によっても,どれだけ多くの遺伝子がその形質発現に関与し,それらの遺伝子が優性か,劣性か,あるいは伴性遺伝するのか,さらにそれらの遺伝子が表現型発現のためにどのように作用しているのかは分からない。
肥満症のようなヒトの複雑な形質に関与する遺伝子を同定するためにいくつかの方法がある。その一つは,ラットやマウスのような動物モデルを用いることである。他の動物で遺伝子が同定されれば,ヒトに同一の遺伝子やその関連遺伝子が存在するかどうか,またそれらがマウスとヒトにおいて同じ方法で働いているのかどうかを知るために,組み換え DNA 技術を用いることができる。
 遺伝子が体重をどのように制御しているのかを理解する上で,画期的な成果が最近マウスの研究から得られた。これまで,体重を制御しているいくつかのマウスの遺伝子が同定,分離,複製,解析されている。マウス突然変異体 obese( ob )と diabetes( db )は共に肥満体である( 右図 )。
遺伝子が体重をどのように制御しているのかを理解する上で,画期的な成果が最近マウスの研究から得られた。これまで,体重を制御しているいくつかのマウスの遺伝子が同定,分離,複製,解析されている。マウス突然変異体 obese( ob )と diabetes( db )は共に肥満体である( 右図 )。
組換え DNA 技術を用い,これらの遺伝子とヒトの相同遺伝子が分離,染色体上の位置づけがなされ,詳細に研究されている。ob 遺伝子は体重制御ホルモンである レプチン leptin ( “thin” という意味のギリシャ語に由来する ) を支配する。
このホルモンは脂肪細胞によって産生される。マウスでは,このホルモンは脂肪細胞から放出され,血中を介して脳に達する。そこでは視床下部の細胞がレプチンに対する細胞表面受容体をもつ。この受容体は db 遺伝子によって支配される。
レプチンが視床下部細胞に結合すると,レプチン受容体が活性化され,遺伝子発現の変化を含む反応が開始する。この反応には食欲抑制ホルモンである glucagon-like protein-1 ( GLP-1 ) の生産を含む。正常な個体では,制御システムが脂肪または筋肉に変換する食物の量を調整する。また,このホルモンはエネルギー消費率を変化させ,比較的狭い範囲に体重が維持されるよう働く。マウスにおける体重の制御には,tubby( tb )のような他の遺伝子も関係している。
レプチンに対するヒト遺伝子( MIM/OMIM 164160 )は,マウス ob 遺伝子と相同で,染色体 7q31.1 に位置する。レプチン受容体の遺伝子( MIM/OMIM 601007 )はマウス db 遺伝子と相同で,染色体 1p31 に位置する。
ほとんどの肥満の人は ob 遺伝子に突然変異をもっているのではなく,レプチンを過剰生産していることが示されている。このような人の場合,受容体,または受容体によって活性化される制御システムに異常があるようである。レプチンとその受容体が体重を制御するのにどのように働いているのかを理解することは,肥満症に対する新薬の開発にもつながる。
アメリカ人は体重を調整する製品に毎年 300 億ドル( 3.8 兆円 )以上費やしているので,レプチンやレプチン関連薬には巨大な市場性がある。バイテク企業はレプチンに関する新薬開発のライセンス料に 2,000 万ドル( 25 億円 )を支払っており,これらの製品は 2 – 3 年のうちに市場に出回ると予想される。
レプチンの遺伝子がある面で体重を制御しているが,ヒトでは,体重のような複雑な表現型を単一遺伝子が制御しているという証拠はほとんど無い。肥満症に直接関連する特定の遺伝子型はほとんど知られていない。体重を制御するヒト遺伝子を探索するために,大規模な,数世代にわたる家族に対して,肥満症と連関する分子マーカーを用いたゲノムレベルの研究が開始されている。
- 肥満症として選抜された 92 家族を用いた研究で,肥満症と第 20 染色体の長腕に位置する 1 個または複数の遺伝子との間に正の相関が認められている。
- また,同様の方法で,体重の制御に関わると考えられる遺伝子座を,第 2 染色体上と第 11 染色体の長腕上に同定したとの報告もある。今後,これらの研究によって,肥満症に含まれる特異的な遺伝子が同定され,体重を制御する分子機構が解明されるだろう。
他の多因子性形質に関する調査
ヒトにおける表現型の多様性の多くは
- 遺伝子と
- 環境要因
との相互作用によって生じる。 これらの形質の多くは多数の遺伝子を含んでいる。ただし, 1 つ 1 つについてみると表現型に対する影響は極めて小さい。これらの各遺伝子は環境と相互作用して,いろいろな表現型を示し,しばしば疾病に関連する遺伝的な要因をぼかしてしまうこともある。後述するように,多因子性形質の遺伝的な要因を決定する手段が考案されている。ここ数年の間に開発された新しいポリジーンの検査方法とヒトゲノムプロジェクトの結果により,遺伝的ならびに環境的要因がどのようにこれらの形質や他の複雑な形質に関与するのかいつかは説明できるだろう。
1. 心疾患には遺伝的ならびに環境的成分が関与する
2 つの因子が心疾患に関与する。すなわち,
- 高血圧( MIM/OMIM 145500 )と
- アテローム性動脈硬化( MIM/OMIM 143890,動脈壁へのプラーク沈着 )
である。両形質は,以下のような環境に大きく影響されるが,すでに明らかにされている遺伝的要因にも依る。
- 遺伝
- 男性であること
- 高血圧
- 高コレステロール値 ( 高 LDL または低 HDL )
- 喫煙
- 肥満症
- 運動不足
- ストレス
高血圧症は血圧が常に 140/90 mmHg 以上の場合である。少なくとも 10 個の遺伝子が血圧を調整するのに関わっている。
それらのほとんどは,腎臓で血中へ再吸収される塩類と水分量を調整するのに作用するようである。それらのひとつが アンギオテンシノジェン angiotensinogen ( AGT ) に対する遺伝子である。
- AGT( MIM/OMIM 106150 )は肝臓で作られ,血中に分泌されるタンパク質である。
- その活性型では,AGT は塩類と水分の保持性を制御する。
- そして,これが血圧を調整することになる。
このタンパク質のいくつかの変異体が,高血圧になりやすい素質に関連があり,初期にほとんど症状がみられないので,俗に silent killer と呼ばれている ( 合衆国の成人人口の 10 – 20% が高血圧症であり,重要な健康問題となっている )。高血圧のすべての型が,必ずしも腎臓において作用する遺伝子によって引き起こされているわけではない。高血圧のまれな型として,常染色体上の優性形質として遺伝する家族性肺高血圧症 ( FPPH ) ( MIM/OMIM 178600 ) が細胞表面受容体の突然変異体として同定されている。発症すると,肺の血管の内皮細胞の過剰生産が起こり,その結果肺動脈の圧が上昇する。
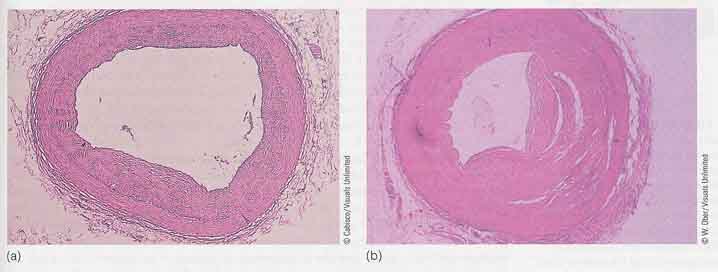
アテローム性動脈硬化は,食事摂取と脂質,とくにコレステロールの生合成,利用,分解との不均衡の結果である。この不均衡により,血管の閉塞や心疾患の原因となる( 上図 )。最も重大な結果は,脳と心臓において血管閉塞が起こった場合で,脳梗塞や心臓発作を引き起こす。
コレステロールは,
肝臓でコレステロールが合成されるので,必ずしも食事に必要な成分ではない。しかし,過剰のコレステロールが動脈プラークに蓄積した場合,心疾患の重要な要因となる。
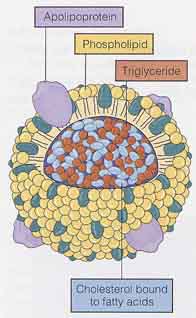 コレステロールのような大きな脂質分子は,血漿に不溶性で,タンパク質とリン脂質の層に包まれ輸送されている。この層の成分は リポタンパク質 lipoprotein として知られている( 右図 )。
コレステロールのような大きな脂質分子は,血漿に不溶性で,タンパク質とリン脂質の層に包まれ輸送されている。この層の成分は リポタンパク質 lipoprotein として知られている( 右図 )。リポタンパク質はその大きさと密度によって分類される。コレステロールは 2 つのタイプのリポタンパク質によって運搬される。すなわち,
- low-density lipoprotein ( LDL,低比重リポタンパク質 )
- LDL は約 45% のコレステロールから成る。
- LDL はコレステロールを肝臓から体の組織に運搬する。
- high-density lipoprotein( HDL,高比重リポタンパク質 )
- HDL は約 20% のコレステロールから成る。
- HDL はコレステロールを肝臓へ輸送する。
である。動脈硬化の危険性は HDL/全コレステロール比と関連する。すなわち,HDL が高いとその危険性は低い。
心疾患の遺伝学的研究は発症する素因となる遺伝子の発見に向けられている。これにより,病気の進行に影響する環境要因 ( 食事や運動など ) を特定でき,また心疾患の危険性をもつ個人を検出できる。
多くの遺伝子が体内のコレステロール濃度を調整する。アポリポタンパク質 ( 細胞受容体に接着するリポタンパク質の一部 ) をコードする遺伝子もその一つである。他の遺伝子はリポタンパク質とその分解酵素に関わる細胞受容体をコードする。受容体遺伝子の突然変異については次に述べる。
常染色体上の優性形質として遺伝する疾病である家族性高コレステロール血症 ( FH ) ( MIM/OMIM 143890 ) は,LDL の摂取を調整する細胞表面受容体の欠損によって起こる。
発症すると ( ヘテロ接合体 ) は血清中のコレステロール濃度が上昇し,たいてい 40 歳から 50 歳の間に冠状動脈疾患を発症する。
 2 種類の突然変異が知られている。
2 種類の突然変異が知られている。- 欠陥受容体と
- 受容体不在
である。いくつかの欠陥受容体の型が知られている。たとえば,
- LDL と認識・結合しない型,
- LDL との結合能力が低下した型,
- LDL と結合・認識するが,細胞に移動できない型など
である。 LDL 受容体をコードする第 19 染色体にある遺伝子の 150 以上の突然変異が確認されている。突然変異のそれぞれが血中の LDL コレステロールの濃度上昇をもたらし,動脈内面にプラークを沈着させる。これが早期の心疾患の原因となる。FH の発症頻度は,
- ヨーロッパ,日本ならびにアメリカ合衆国 – 約 1/500
- ケベック – 1/122
- 南アフリカ – 1/71
である。平均頻度 1/500 ということは,この病気がヒトにおける最も一般的な突然変異の 1 つによるものであることを示唆する。
脂質代謝を調節する他の遺伝子における突然変異も心疾患の原因である。アテローム性動脈疾患感受性 ( ATHS ) の遺伝子が第 19 染色体の LDL 受容体の遺伝子座近くにマップされている。ATHS ( MIM/OMIM 108725 ) により,血中 LDL 濃度が上昇,HDL 濃度が減少して,心臓発作の危険性が 3 倍に高まる。
2. 皮膚色は多因子性形質である
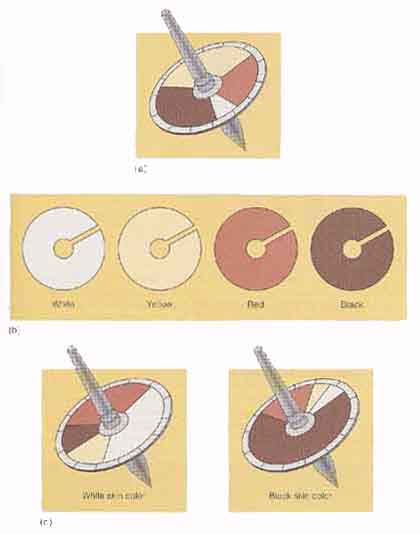
 Davenport 夫妻がヒトのいくつかの形質が多因子性であるという見解を初めて検証した。1910 年から 1914 年にかけて,バミューダ諸島や西インド諸島における黒人と白人の夫婦について皮膚色の情報を収集した。皮膚色は遺伝ならびに環境要因によって制御されており,多因子性形質である。陽に当たると日焼けして,遺伝子型の差異がぼやけてしまうので,Davenport 夫妻は上腕部の皮膚色を測定した。
Davenport 夫妻がヒトのいくつかの形質が多因子性であるという見解を初めて検証した。1910 年から 1914 年にかけて,バミューダ諸島や西インド諸島における黒人と白人の夫婦について皮膚色の情報を収集した。皮膚色は遺伝ならびに環境要因によって制御されており,多因子性形質である。陽に当たると日焼けして,遺伝子型の差異がぼやけてしまうので,Davenport 夫妻は上腕部の皮膚色を測定した。皮膚色を測定するために,彼らは色領域が変更できる円盤を組み込んだ独楽 ( コマ ) を用いた( 右上図 )。色領域は,黒,白,赤,ならびに黄色で,コマを回した時のブレンド色が皮膚色と一致するように,色領域を変更して,各色領域の割合が決定できるようになっている。Davenport 夫妻は,皮膚色と一致した場合の黒色領域の大きさを測定し,調査対象者を 0 から 4 までの 5 段階に分類した。
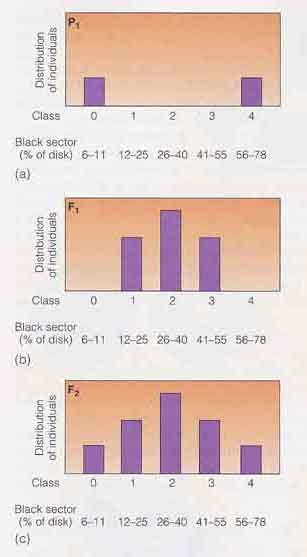
この調査の結果により,ポリジーン形質のいくつかの特徴が明らかとなった。
- 子供( F1 )は両親の皮膚色の値の中間値をとる。
- F 2 世代では,少数の子供が祖父母の一方と同じように白色であり,また少数が祖父母の他方と同じ黒色であった。
- そして,ほとんどの子供たちは両者の中間の皮膚色を示した( 右図 )。
F2 個体が 5 段階 ( クラス ) の表現型群に分類できたので,Davenport 夫妻は皮膚色には 2 個の遺伝子が関与していると仮説を立てた。F2 の各表現型群は,遺伝子の分離と組み合わせによって生じた遺伝子型を表わすと考えられた。この結果を説明するために,これらの遺伝子を仮に A と B としよう。クラス 0 は最も明るい皮膚色をもち,遺伝子型は aabb で示される。クラス 1 は遺伝子型 Aabb または aaBb をもち,そして最も濃い皮膚色で優性遺伝子がホモ ( AABB ) であるクラス 5 まで以後同様である。
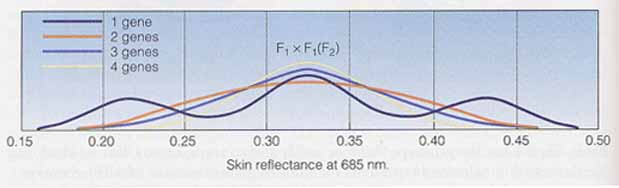
その後,皮膚表面の光反射を測定できる装置を用いた他の研究者によって,皮膚色は実際 2 対以上の遺伝子によって支配されることが示されている。3 個または 4 個の遺伝子を想定したモデルが最もデータと一致するようである ( 下図 ) 。
3. 知能と知能指数 ( IQ ) : 相関はあるのか?
20 世紀初頭に,フランスの心理学者 Alfred Binet は知能を測定するために,心理学的な方法を採用した。特殊教育を必要とする子供を識別するために,理解,選別ならびに訂正などの基礎的な心理作用に関連する段階別課題を開発した。子供たちはそれぞれ最も簡単な課題から始め,課題が難しすぎるようになるまで順々に進めていく。達成された最後の課題に割り振られた年齢が,その子供の精神年齢とされ,知能年齢は実年齢から精神年齢を引いて計算される。Binet テストは今日用いられている Stanford-Binet 知能テストの基礎となっている。
心理学者 Wilhelm Stern は精神年齢を実年齢に対する比率で示すことを提唱し,知能指数 intelligence quotient ( IQ ) として知られるようになった。
7 歳の子供が,7 歳用の課題をうまく解いて,8 歳用の課題を達成できなかったとすると,精神年齢は 7 が割り振られる。この子供の IQ を求めるには,精神年齢を実年齢で割る:すなわち,精神年齢 7 ÷ 実年齢 7 = 1.0。小数点をなくすために,商を 100 倍する ( 1.0 × 100 = 100 ) 。この子供の IQ は 100 と求められる。
IQ テストや他の方法で知能を正確に測定できるかどうかを明確にするためには,身長,体重や指紋を測定したような方法で客観的に測定できるよう,まず知能を定義しなければならない。推理能力,計算能力,表現能力,判断能力,解決能力ならびに創造能力などの特性が知能の重要な構成要素であるとよく言われている。
残念ながら,これらの特性のどれもが IQ テストで測定できるという証拠はなく,知能の上記構成要素を定量化する客観的な方法もない。
知能に関連する精神機能には多種多様な神経過程が含まれるだろう。しかし,精神生物学の急速な進展にもかかわらず,これらの過程のいずれの生物学的な基礎についても情報がほとんど無い。
しかし,IQ 測定で得られた数字は遺伝的な要素と考えられる。この証拠は次の観察から得られている。
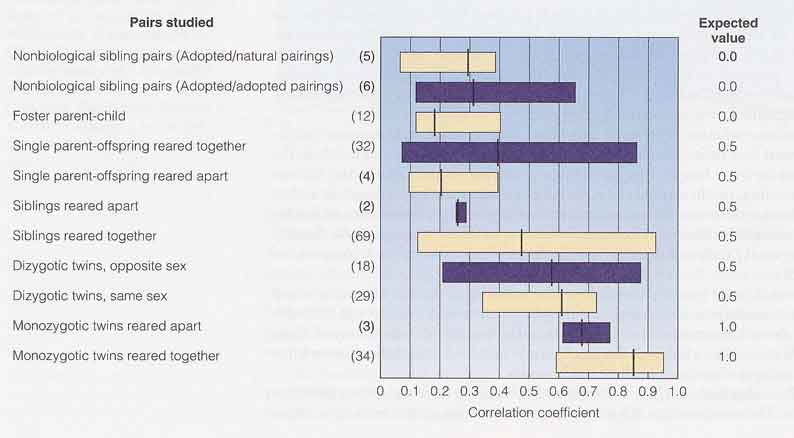
4. 知能を制御する遺伝子の探索
肥満症の項で述べたように,遺伝率を研究しても,知能に関わる遺伝子の数や局在,遺伝子の作用についての情報は得られない。遺伝子本体について知るために,IQ に勝る知能についての拡大された定義付けがされ,すなわち一般認識能力 general cognitive ability と定義された認識ならびに知的能力には多くの遺伝子が関与していると考えられている。
知能に関連する遺伝子を同定することを目的として,ハエ Drosophila やマウスの動物モデルが,学習,記憶,空間認知を制御する遺伝子の同定に用いられている。これらの遺伝子の染色体上の位置が決定され,その特性について情報が得られれば,ヒトにおいて相同遺伝子の同定,位置決め,分離が行われるだろう。
2 つ目の研究方法は,分子遺伝学と組換え DNA 技術を用いて,読解力や IQ のような特別なポリジーン形質に影響する遺伝子を同定することである。これらのポリジーンは 量的形質座位 quantitative trait loci ( QTLs ) として知られている。
QTLs の探索により,これまで 失読症 dyslexia の関連遺伝子が第 6 染色体 ( MIM/OMIM 600202 ) と第 15 染色体上にあることが判明している。また最近,一般認識能力に関連する遺伝子が第 6 染色体上に同定されている。
行動遺伝学,双子研究,分子遺伝学を組み合わせた研究方法により,近い将来,認識能力に関連するより多くの遺伝子が同定されるだろう。
ヒトゲノムプロジェクトの結果が解析されるにつれ,より高度な精神過程に含まれる遺伝子の数や作用を決定することが容易になり,知能の遺伝学に多くの情報をもたらすだろう。
そのような遺伝子が同定,分離されれば,今度は特定の遺伝子型をもつ個人に対する遺伝的差別や知能を増強するための遺伝子治療の可能性などが危惧される。
最初に戻る
メニューのページへ戻る February 07, 2020