細胞呼吸
| このページの内容 |
細胞呼吸は,グルコースのような栄養分子を酸化して,二酸化炭素と水に分解する過程である。放出されるエネルギーは ATP の形で捕捉され,細胞のすべての活動に利用される。
この過程は,以下の 2 段階で起こる:
- 解糖 glycolysis, グルコースがピルビン酸は分解される。
- ピルビン酸の完全な 酸化 によって,二酸化炭素と水に分解される。
真核生物では,解糖 は細胞質で起こる。次の過程は ミトコンドリア で起こる。
ミトコンドリア
ミトコンドリアは膜に包まれた細胞小器官で,ほとんどの真核細胞の細胞質全体に分布している。その主な機能は栄養分子のポテンシャル・エネルギーを ATP に変換することである。 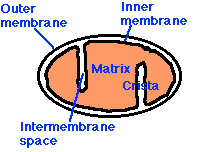 ミトコンドリアは,以下の構造物からなる:
ミトコンドリアは,以下の構造物からなる:
- 外膜 outer membrane – すべての構造を包む。
- 内膜 inner membrane – 液体で満たされた基質を包む。
- 両者の空間が 膜間腔 である。
- クリステ cristae – 内膜は棚状に折り畳まれ,クリステを形成し,基質中に突出している。
- 少数 ( 5 – 10 個程度 ) の環状 DNA 分子
外膜
外膜は多くの内在性タンパク質複合体を含み,ミトコンドリアに出入りする多種多様な分子やイオンが通過するチャンネルを形成している。
内膜
内膜は,以下の 5 種類の内在性タンパク質複合体を含む:
- NADH デヒドロゲナーゼ
- コハク酸デヒドロゲナーゼ
- シトクロム c 還元酵素 ( シトクロム b-c1 複合体とも呼ばれる )
- シトクロム c 酸化酵素
- ATP シンターゼ
基質
内膜に囲まれた基質にはクエン酸回路で働く可溶性酵素や,脂肪酸の b 酸化にかかわる諸酵素が含まれている。
ここでピルビン酸は,
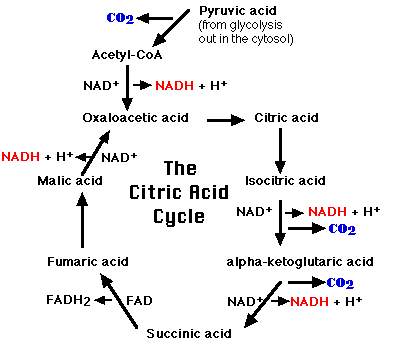
- NAD+ によって酸化され, NADH + H+ を生成する。
- カルボキシル基が除去され,以下の分子を生成する:
- 二酸化炭素 ( CO2 ) と
- コエンザイム A の SH 基に酢酸がチオエステル結合して,を生成する。
クエン酸回路
- アセチル-CoA は オキサロ酢酸 と縮合して,
- クエン酸が生成する ( 右図には一連の酵素反応を示した ) 。
- 最後の段階で,リンゴ酸は NAD+を補酵素とするリンゴ酸デヒドロゲナーゼの作用で再びオキサロ酢酸を生成することとなる。こうしてクエン酸回路が一回転する。
要約:
- ミトコンドリアに入ったピルビン酸に存在する炭素原子 3 個は二酸化炭素 ( CO2 ) として放出される。
- 4 つの反応で,電子対が ( 2e– ) 除去され,NAD+ に引き渡され,それが還元されて NADH + H+ となる。
- 1 つの反応で,電子対がコハク酸から除去され, FAD が還元されて FADH2 となる。
NADH と FADH2 の電子は次に述べる呼吸鎖の経路に引き渡される。
呼吸鎖
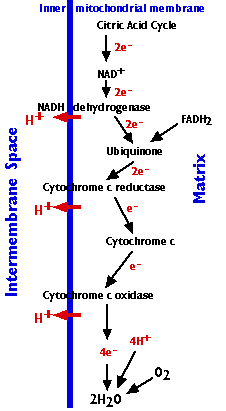 呼吸鎖は,次の 3 つの内在性タンパク質複合体からなる:
呼吸鎖は,次の 3 つの内在性タンパク質複合体からなる:
- NADH デヒドロゲナーゼ 複合体
- シトクロム c 還元酵素 複合体
- シトクロム c 酸化酵素 複合体
ならびに自由に拡散できる 2 つの分子が含まれる。
- ユビキノン ubiquinone
- シトクロム c cytochrome c
これが複合体から複合体へ電子を送り出す。
呼吸鎖:
- NADH ( ならびにFADH2 ) から電子が 酸素 分子に段階的に引き渡され,水 ( H2O ) となる。( シトクロム c は一度に 1 個の電子を引き渡すことができる。したがって,シトクロム c 酸化酵素は,酸素と反応するために,電子が 4 個蓄積するまで待機している。)
- 基質 から 膜間腔 へ陽子 ( H+ ) が次々に引き渡されることによって放出されるエネルギーを利用している。
- 酸素を還元して水になるに必要な 4 個の電子が呼吸鎖を通過する時には,約 20 個の陽子が膜間腔へ移送される。
- このような能動輸送によって内膜を超えて形成された陽子の濃度傾斜は,小型のバッテリーに匹敵する。
- 陽子はこの濃度傾斜の下で,別の内膜の内在性タンパク質複合体,すなわち ATP シンターゼ 複合体を利用して基質に再度還流する。
ミトコンドリアにおける化学浸透性
呼吸鎖電子伝達反応が進行する際に,H+ が膜を横切って一方から他方へ定方向的に輸送され,(2) ミトコンドリアの基質から膜間腔へ濃度傾斜に逆らって陽子 ( H+ ) を送るために,電子が NADH から酸素への濃度傾斜を通過する時に放出されるエネルギーが,呼吸鎖の 3 つの酵素複合体によって利用される。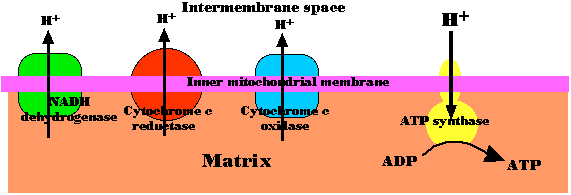
その結果として生じる 陽子 ( H+ ) の偏在が ATP 合成のエネルギー源となる。この過程は 化学浸透 と呼ばれ,1966 年ミッチェル P. Mitchell によって提唱された。
ATP のモル数
ほとんどの ATP は,ミトコンドリア内膜を横切って形成された陽子の濃度傾斜によって生成される。ATP シンターゼを通って陽子が戻る時に,電子対当たり 3 モルの ATP ( ただし, FADH2 に結合したものでは 2 モルの ATP )が合成される。
グルコース 1 モルから 12 対の電子が遊離するので,
- 10 x NAD+ ( 10 x 3 = 30 )
- 2 x FADH2 ( 2 x 2 = 4 )
したがって,34 モルの ATP が生成される。
この他に 4 モルの ATP が生成されるので,合計 38 モルとなる。
その内容は:
- クエン酸回路の 1 ステップで ( a-ケトグルタール酸からコハク酸への変換時に ) グルコース 1 モル当たり 2 モルの ATP が生成する。
- 解糖 系でグルコース 1 モル当たり 2 モルの ATP が生成する。
しかし,ミトコンドリアには,必要な分子やイオンの能動輸送など,他にもたくさんの機能があり,陽子の濃度勾配のエネルギーはこれらにも利用される。
また,NADH は多くの細胞反応の還元剤としても用いられている。
したがって,実際の ATP 量は細胞の状態によって変動するる。おそらく,30 モルを超えることはないだろう。
ミトコンドリア DNA (mtDNA )
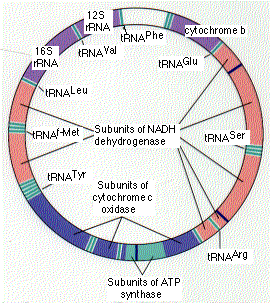
ヒトの ミトコンドリア mitochondrion は, 5 – 10 個の環状 DNA 分子を含んでいる。各 DNA 分子は,以下をコード化する 37 個の遺伝子を含む,16,569 個の塩基対から成る:
- 2 種類のリボソーム RNA ( rRNA )
- 22 種類の 転移 RNA ( tRNA ) ( 少なくとも各アミノ酸に 1 個が対応している )
- 13 種類の ポリペプチド
rRNA と tRNA 分子は 13 種類のポリペプチドを合成する機構に利用される。
この 13 種類のポリペプチドはミトコンドリアの内膜にあるタンパク質複合体のサブユニットで, NADH デヒドロゲナーゼ,シトクロム c 酸化酵素 ならびに ATP シンターゼ のサブユニットも含む。しかし,これらのタンパク質複合体のそれぞれには,核の遺伝子にコード化されたサブユニットも必要である。すなわち,それらは細胞質で合成された後,ミトコンドリアへ移送されるものである
mtDNA の突然変異は遺伝病の原因となる
ミトコンドリア遺伝子の突然変異によって多くの疾病が起こる。たとえば,
- シトクロム b
- 12S rRNA
- ATP シンターゼ
- NADH デヒドロゲナーゼのサブユニット
- 数種類の tRNA 遺伝子
様々な器官が影響されるが,脳および筋肉の遺伝病が最も一般的なものである。これは,おそらくこれらの両器官のエネルギーの大きな需要を反映しているものと思われる。
これらの障害のいくつかは生殖細胞を介して受け継がれる。精子のミトコンドリアは受精卵の中に残らないので,すべての場合の突然変異遺伝子は母親から受け継いだことになる。他の遺伝病は 体細胞 に起こった突然変異が原因となる。
例: 運動不耐性
運動不耐性 ( 筋肉が容易に疲労してしまう ) に苦しむ多くのヒトは,シトクロム b 遺伝子の突然変異をもつ。奇妙にも,彼らの筋肉中のミトコンドリアだけ突然変異を示す。他の器官のミトコンドリア正常である。おそらく,胚発生の極めて早期にある細胞で,シトクロム b 遺伝子に突伝変異が起こり,この細胞が筋肉に発生する運命にあったものと考えられる。
なぜミトコンドリアは自らのゲノムを持っているのだろうか?
ミトコンドリアの遺伝システムの特徴の多くは,細菌のような前核生物にみられるものに近い。このことは,ミトコンドリアが進化的に前核生物の子孫であるとする説を支持する。
すなわち,ミトコンドリアは地球上における生命誕生の初期に,真核細胞の先祖との 内部共生的 endosymbiotic な関係を築いた前核生物の子孫と考えられている。しかし,ミトコンドリアの機能に必要な遺伝子の多くはその後核ゲノムに移ったと考えられている。
最近,Rickettsia prowazekii のゲノム の塩基配列決定がなされ,その多くの遺伝子がミトコンドリアにみられるものと極めて近い関係にあることが分かってきた。おそらく,リケッチアの仲間が真核生物のミトコンドリアになった内部共生者と最も近縁の生きた子孫であろう。
| 最初に戻る |
January 31, 2020