| このページの内容 |
生殖細胞の不思議?!
生殖とは
有性生殖とは 2 種類の配偶子の融合による新しい個体の形成である。ヒトおよび大多数の真核生物には, 2 種類の配偶子がある。
- 構造が異なる ( 異形配偶子 )
- 異なる個体 ( 両親 ) から由来する
配偶子の特徴
- 会合し,融合するための運動性
- 初期発生のための栄養素
動物では,これらの相反する要求が,異形配偶子によって満たされる。
- 小型で,運動性をもつ精子
- 栄養素を含有する卵子
生殖システム
ヒトの生殖は下記システムの統合された働きによって営まれている。
生殖細胞の不思議?!
- 1 個の精子と 1 個の卵子が融合して,両親と子供との間に遺伝的な関連が生じる。
- 将来の精子または卵子となるよう運命づけられた細胞は胎児期の早期に明確に分離される。
例 1:
妊娠の 15 週までに,女の赤ちゃんは将来卵子となる細胞を別に確保している ( 事実,これらの細胞は 減数分裂 meiotic division の最終段階に進行している ) 。
例 2:
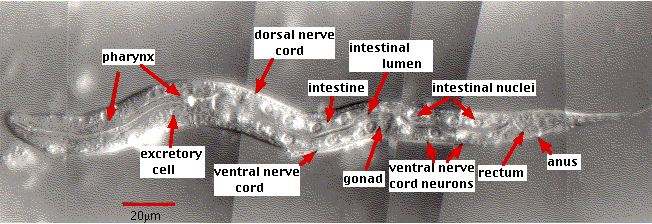
Caenorhabditis elegans は顕微鏡でないと見えない大きさの ( 約 1 mm ) 線虫で,通常は土壌中に生息する。すべての動物と同様に,この線虫も受精卵 ( 接合子 ) から一生が始まる。受精後,有糸分裂 mitotic division によって,
- 新たに付加した線虫の 556 個の細胞と,その後に,
- 成体になって,959 個の体細胞と,種々の数の生殖細胞が生じる。
この線虫は,
- 透明で,体が観察しやすく [ 顕微鏡写真 ] ,また
- 分化のパターンが極めて厳格である。
{kind=link}
したがって,この動物ではすべての体細胞の細胞系譜を追跡することが可能である。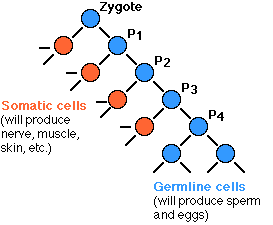
接合子の最初の 4 回の有糸分裂で生じた 16 細胞の運命が完全に予測できることが判明している。
もちろん,接合子は 全能性 totipotent をもっている。このことは,この動物のすべての細胞,すなわち生殖細胞と体細胞を形成する能力を持っていることを意味する。
最初の 4 回の有糸分裂で, 1 個の娘細胞 ( 赤 ) はその全能性を失う。そして,この細胞のすべての後継細胞がこの動物の体細胞組織を形成するように限定されることになる。図中,青で示した 2 番目の娘細胞はその全能性を保有している ( P1, P2, P3, ならびに P4 ) 。
5 回目の有糸分裂で,両方の娘細胞が全能性を保有したまま,時期が来ると精子や卵子を形成する。
ワイスマンの生殖質理論
100 年以上も前に,ドイツの動物学者ワイスマン Weismann は,体細胞系 soma と生殖細胞系 germline を区別し,動物は 2 種類の細胞から成ると考えた:
- 体を構成する細胞 ( 体細胞系 somaplasm ), そしてこれには
- 配偶子をつくる細胞 ( 生殖細胞系 germplasm ) が含まれる。
各世代で,接合子から発生する胚は,次世代のために生殖細胞質を保存するだけではなく,生物の体を構成する細胞も生産する。
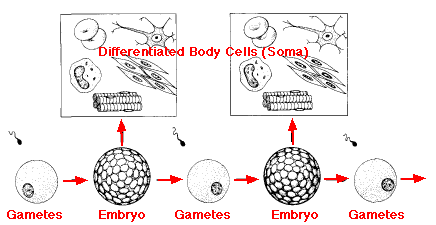
| ワイスマンのいう germplasm,すなわち配偶子とそれらを形成する細胞( 生殖細胞系 )においてのみ テロメラーゼ telomerase 活性が高く維持されている。 これらの細胞では染色体の長さが永久に維持され,不死とされている。これに対して,体細胞系はテロメラーゼを生産するのを止め,有糸分裂毎に染色体の先端部が消失して,結果的に死滅する。 |
ワイスマンは,種の保存を永続させるために生殖細胞系が重要であり,体細胞系 somaplasm は 生殖細胞系 germplasm を保護する器であり,次世代を形成するために,生殖細胞系 が保護され,養われ,異性の 生殖細胞系 まで運ばれるように面倒を見ていると考えた。
どのように生殖細胞系列と体細胞が決定されるのだろうか?
- 胚の初期のどの細胞が精子や卵子を形成していく運命にあるのか,すなわち 生殖細胞系列 germline に入るのはどの細胞か?
- 動物の体の組織になるのはどれか?
昆虫
昆虫では,卵の一端に母親によって挿入された特別の細胞質によって制御されている。
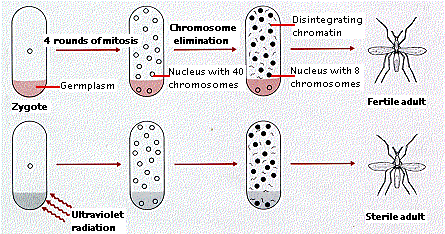
タマバエ ( 昆虫 ) 卵の 4 回目の有糸分裂で, 16 個の核のうちの 2 個が卵の一端に少量の細胞質と共に押しやられる。 5 回目の有糸分裂時に,これらの 2 個の核が正常に分割し,この種の完全な染色体数 ( 2n = 40 ) をもつ娘細胞を形成する。
しかし,その他の核ではそうならない。それぞれで分裂後期に達した時に,40 本の染色体 ( 二分染色体 ) のうち 8 本だけが分離して,紡錘極に移動する。残りの 32 本の染色体は赤道面に留まり,最終的には崩壊していく。
- この 2 個の“正常な”核の後継細胞が最終的に精子や卵子を形成するよう,すなわち生殖系列に分化する。
- 残りの核の後継細胞は,著しく染色体数が減少して,この昆虫の体を構成する ( 上記以外の ) すべての組織,すなわち体細胞を形成する。
右図の上段はタマバエにおける正常な発生を示す。配偶子は,完全な染色体数をもつ 2 個の核からすべて生じる。それらは細胞質の特別な領域 ( ここでは生殖細胞質 “germplasm” とよぶ ) に区分されている。残りの核は 32 本の染色体を失い,最終的に体細胞を形成する。
下段は生殖細胞質を破壊することによって,そこに移動した核から染色体を除去した場合を示す。この場合,タマバエは不妊となるが,他の部分は全く正常である。
哺乳動物
哺乳動物では ( 少なくともマウスでは – Saitou et al, 2002 年 7 月 18 日号の Nature 参照 ) , 生殖細胞になる決定は本来備わっているものではなく,初期胚発生中の 細胞と細胞の信号伝達 の結果である。
将来の生殖細胞が確保され始めるのは 原腸形成 gastrulation が始まる時 ( マウスでは受精後 6 日目 ) のようである。この時期は,中胚葉が形成されている時でもある。
以下のように進行する:
ステップ 1
将来の 胚体外膜 extraembryonic membranes となる 外胚葉 が 骨形成タンパク質 bone morphogenic protein,たとえば Bmp-4 を分泌する。これは,発育中の中胚葉に信号を与え,以下の将来的にいずれかを形成するようにさせる。
- 胚体外膜 ( 羊膜と尿膜 ) の中胚葉, または
- 始原生殖細胞 primordial germ cell ( PGC )
ステップ 2
これらの細胞の部分集合 ( 集合体の中央で ) 以下の発現を示すようになる:
- 遺伝子 fragilis によってコードされている膜貫通型タンパク質;
- stella ( またはPgc7 )として知られている DNA-結合タンパク質 ( 転写調節因子 か? )
( Stella/Pgc7 は接合子にも存在し,胚性幹細胞 embryonic stem cell にも認められている。) - 胎児の体組織や胚体外膜の発生に必要な遺伝子 ( たとえば, Hoxb-1 のようなホメオボックス遺伝子 )を抑制する。
ステップ 3
PGCs が将来生殖腺 ( 卵巣 または 精巣 ) を形成すべく,発育中の胎子の特定の部位に移動する。
ワイスマン理論の例外
生殖細胞と体細胞は 動物 においてのみ区別される。
植物 では,配偶子となる細胞も体細胞組織から生じる。
微生物 では,すべての機能が 1 つの細胞に統合されている。
( しかし,繊毛をもつ原生動物 テトラヒメナ Tetrahymena の様なある種の単細胞動物では,小核 micronuclei 内に完全なゲノムをもち,これが次世代に受け継がれる。一方,大核 macronucleus の遺伝子は受け継がれない。このように,これらの生物では生殖細胞と体細胞の区別らしきものが存在する。)
体細胞と生殖細胞の突然変異
生殖細胞の突然変異の頻度
[ 突然変異の頻度についてはこちら ]
| 最初に戻る |
| メニューのページへ戻る |
February 06, 2020