内部共生
| このページの内容 |
内部共生説の概要は,以下の通り。
- 真核生物の ミトコンドリア mitochondria はリケッチアなどの好気性菌が宿主細胞に住み着いて進化した。
- 葉緑体 は光合成細菌やラン藻の祖先が,もともとは葉緑体をもたなかった宿主細胞と共生した結果生じた。
- 真核生物の 繊毛 ならびに鞭毛は内部共生したスピロヘータから生じた。真核生物の繊毛や鞭毛が発達した基底小体から紡錘体が形成されて, 有糸分裂 が可能となった。
内部共生を支持する根拠
それを支持する根拠としては,ミトコンドリア内のタンパク質合成系が,ミトコンドリアの小さなリボソーム上で行われ,細菌のタンパク質合成を阻害する抗生物質クロラムフェニコールやストレプトマイシンに感受性があるなど真核細胞細胞質の大きなリボソーム上で行われるタンパク質合成と異なっていること,また一部のアミノ酸に対する遺伝情報のコドンがミトコンドリアの DNA と核 DNA とで違った組合せのものを使っていること などである。しかしミトコンドリアが自己増殖系としては全く不完全で,核遺伝子に大部分を依存している点の説明がついていない。
以下に,根拠をまとめる:
- ミトコンドリアと葉緑体は前から存在している祖先種 ( 前ミトコンドリアと前葉緑体 ) から生じた。細胞の核ゲノムにはミトコンドリアと葉緑体が利用する数種のタンパク質がコード化されているだけで,細胞ゲノムだけではそれらを複製できない。
- ミトコンドリアと葉緑体は自身の ゲノム を有し,前核生物のそれに似ており, ( 真核生物の ) 核のゲノムとは異なる。
- ゲノムは 環状 DNA 分子からなる。
- DNA に結合する ヒストン をもたない。
- ミトコンドリアと葉緑体は自身のタンパク質合成をもっている。それは前核生物の仕組みに似ているが,真核生物のタンパク質合成と異なっている。
- それらの リボソーム RNA ( rRNA ) とリボソームの構造は前核生物のそれに似ているが,真核生物のものとは異なる。
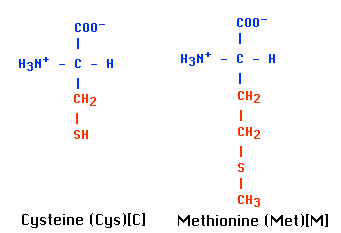
- それらの転写の開始コドンは,細菌のように常に fMet である( 真核生物のように メチオニン [ Met ] ではない ) 。
- 細菌におけるタンパク質合成を抑止するよう作用する多くの抗生物質 ( たとえば,ストレプトマイシン ) は,ミトコンドリアと葉緑体内のタンパク質合成も抑止する。これらの抗生物質は,真核生物の細胞質内でのタンパク質合成には影響しない。
- 逆に言えば,真核生物のリボソームによるタンパク質合成の抑制剤 ( たとえば,ジフテリア毒素 ) は細菌のタンパク質合成もミトコンドリアと葉緑体内のタンパク質合成にも目立った影響は示さない。
- 抗生物質 リファンピシン は細菌の RNA ポリメラーゼを抑制し,ミトコンドリアの RNA ポリメラーゼも抑制する。しかし,真核生物の核内の RNA ポリメラーゼには影響がない。
{kind=link}
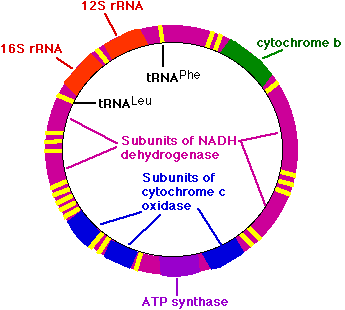
ミトコンドリアのゲノム
ヒトのミトコンドリアのゲノムは 16,569 塩基対の環状 DNA からなる。これらは,以下のものがコード化されている:
- 2 種類の リボソーム RNA ( rRNA ) 分子
- 22 種類の 転移 RNA ( tRNA ) 分子 ( 図には黄色の線で示してある。それらの 2 つにだけはラベルが付いている )
- 13 種類の ポリペプチド
13 種類の ポリペプチド は ミトコンドリア内膜 に埋め込まれているタンパク質複合体を構築するのに関わっている。
- ミトコンドリアの NADH デヒドロゲナーゼ を構成する 7 個のサブユニット
- シトクロム c 酸化酵素 の 3 個のサブユニット
- ATP 合成 の 2 個のサブユニット
- シトクロム b
これらのすべての遺伝子産物はミトコンドリア内で用いられるが,ミトコンドリアには核の遺伝子によってコード化されているタンパク質も必要である。そのタンパク質 ( たとえば,ミトコンドリア内で用いられる シトクロム c, RNA ポリメラーゼや DNAポリメラーゼ ) は,細胞質内で合成され,その後ミトコンドリアに輸送される。
葉緑体のゲノム
葉緑体のゲノムは ゼニゴケ ( ゼニゴケ類, 蘚苔植物の 1 種 ) で判明している。これは 121,024 塩基対の環状 DNA 分子をもつ。以下のものを含む,128 個の遺伝子からなる。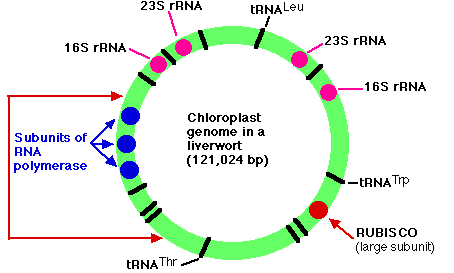
- 葉緑体によって用いられる リボソーム RNA ( rRNA ) の 4 つのサブユニット ( 23S, 16S, 4.5S, ならびに 5S ) それぞれをコード化している重複遺伝子
- 翻訳 に用いられているすべての転移 RNA ( tRNA ) をコード化している 37 個の遺伝子。それらのいくつかを図に黒線で示した ( 一部にはラベルを付けてある ) 。
- 葉緑体内の 転写 に用いられている RNA ポリメラーゼのサブユニットのいくつかをコード化している 4 個の遺伝子( それらの3つを青で示した )。
- 酵素 RUBISCO ( ribulose bisphosphate carboxylase oxygenase, リブロース二リン酸カルボキシラーゼ酸素添加酵素 ) の大サブユニットをコード化する遺伝子。
- 光化学系 I と II の構成分の 9 個の遺伝子
- 葉緑体 ATP シンターゼの一部をコード化する 6 個の遺伝子
- 葉緑体リボソームを構成する約 60 種類のタンパク質のうち 19 種類の遺伝子
すべての遺伝子産物は葉緑体で用いられるが,すべての葉緑体構造に関わるタンパク質は,
- 核の遺伝子によってコード化されている。
- 細胞質で翻訳される。そして,
- 葉緑体に運ばれる。
たとえば,RUBISCO はリブロース二リン酸に CO2 を付加し,カルヴィン回路を開始させる酵素である。 2 つのサブユニットの重複コピーからなる:
- 葉緑体のゲノム内にコード化され,葉緑体内で合成される大サブユニット,ならびに
- 核のゲノムでコード化され,細胞質のリボソームで合成される小サブユニット。その後,葉緑体に輸送される。
真核生物の葉緑体の進化は,シアノバクテリアの内部共生によってそれぞれ別々に 3 回起こったと考えられている。すなわち:
- 緑藻類と植物
- 紅藻
- glaucophytes; 単細胞性藻類の仲間
二次内部共生: 真核生物を飲み込んだ真核生物
核変形体
従属栄養で,光合成による真核生物が進化したとすると,前者は繰り返し後者を飲み込み,独立栄養の仕組みを活用しようとしたと考えられる。今日見られる動物のこの目的で藻類を飲み込んでいる。 たいてい,これらの双利共生関係にあるパートナーは別々に発育する。
しかし,ある藻類の 葉緑体 は上述したような一次内部共生でシアノバクテリア ( 前核生物 ) を飲み込んで生じたのではなく, 光合成による真核生物 を飲み込むことによって生じたと考えられる証拠が報告されている。これを, 二次内部共生 と呼んでいる。 ずっと以前に起こったので,内部共生体は宿主から離れて養われることがなかった。
以下の 2 つの群で,内部共生体が核の痕跡を保持していること ( これを核変形体 nucleomorph と呼ぶ ) からも,内部共生体が真核生物の性質を持つことがうかがわれる。
- 単細胞性の, 運動性をもつ藻類 ( cryptomonads ) の群は, 飲食作用 によって 紅藻類 を飲み込んだ光合成をしない鞭毛をもつ真核生物 ( たとえば,原生動物 ) の進化の結果のようである。
- 他の単細胞性の藻類群 ( chlorarachniophytes ) は 緑藻類 を飲み込んだ鞭毛をもつ原生動物の子孫と考えられる。
2 つの例では,運動性をもち,独立栄養の細胞が,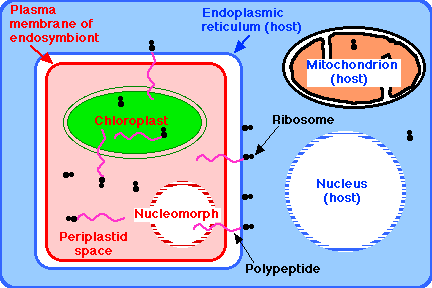
- それ自身の核
- それ自身のミトコンドリア
- それ自身の小胞体をもち,これが以下をもつ内部共生体を包含している:
- 自身の細胞膜
- 自身の細胞質,
- 自身のリボソーム
- 自身の葉緑体, ならびに
- その 核変形体 – 本来の核の痕跡程度に変化しているが,なおも,
- 核膜孔複合体 をもつ核膜によって囲まれ,
- 分子量は小さいが,機能的なゲノムを含む。
4 つのゲノムをもつ Guillardia theta
地中植物 Guillardia theta は 4 つのゲノムをもつ:
- それ自身の核のゲノム: 最も大きく ~350×109 塩基対 ( bp )
- ミトコンドリアのゲノム:48,000 bp
- 内部の葉緑体のゲノム:121,000 bp
- 核変形体のゲノム:551,264 bp
Susan Douglas と共同研究者 ( 2001 年 4 月 26 日号 Nature ) が核変形体のゲノムの塩基配列を報告した。
それには,以下の情報が乗っている 3 本の小さな染色体が含まれる:
- メッセンジャー RNA 以外の RNA ( rRNA, tRNA, snRNA ) の 47 個の遺伝子
- メッセンジャー RNA の 464 個の遺伝子; これは,以下のタンパク質をコード化している:
- それ自身のリボソームの 65 種のタンパク質
- その葉緑体の 30 種のタンパク質 ( 必要となる何百種のうちの一部 )
- 核変形体内に必要な種々のタンパク質,これらには以下が含まれる:
- DNA ライセンシング因子
- ヒストン
- DNA 複製に必要なタンパク質 (だたし, DNA ポリメラーゼに関する遺伝子はない。これは宿主のリボソームで翻訳され,合成されたものが利用される。)
3 本の染色体には遺伝子がぎっしり乗っている。事実,それらの 44 個がお互いに重なり合っている。17 個の遺伝子にだけに イントロン が含まれるが,これらも非常に小さい。
地中植物 Guillardia theta におけるゲノムの相互関係
何百万年もの進化の過程で,4 つのゲノム間の相互関係は複雑であるが,正確に組み合わせられた配列となっている。例えば:
- 葉緑体は 3 つのゲノム ( すなわち,自分自身,核変形体,ならびに宿主のゲノム ) によって合成されるタンパク質を必要とする。
- 核変形体のゲノムは,通常の代謝機能に必要な酵素をコード化している遺伝子の ( 1 つを除く ) すべてが処分されてしまっている。すなわち,内部共生体は今宿主の核によってコード化されている遺伝子に依存しているのである。
- 核変形体自身は宿主の核に属する遺伝子 ( たとえば,DNA ポリメラーゼの遺伝子 ) に依存している。
アピコプラスト Apicoplast
アピコプラスト apicoplast ( “apicomplexan plastid” を短縮したもの ) は,apicomplexan 原生生物で発見された単独の細胞小器官である。これは, Plasmodium falciparum や トキソプラズマ Toxoplasma gondii のような “胞子虫” である。
その特徴は:
- この原生動物は,アピコプラストなしでは生存できない。
- 4 枚の膜に入っている。
- 環状 DNA ( 35,000 塩基対 ) からなるそれ自身のゲノムを含む。これは。以下のものをコード化する:
- 約 30 種のタンパク質
- 必要なすべての転移 tRNA といくつかの他の RNA
- 限られた機能を持つことが判明している。それらは,以下を含む:
- 明らかに,30 種のタンパク質だけではこれらの機能を果たすのには不十分である。したがって,アピコプラストは約 500 種の核がコード化しているタンパク質を細胞質から取り込んでいる。
アピコプラストの DNA の塩基配列は,緑藻類の 葉緑体 のゲノムに非常に近い ( したがって,すべての緑色植物のそれと類似している ) 。
これはすべて,アピコプラストが真核生物の先祖種がただ一つの葉緑体を備えた単細胞性の緑藻類を飲み込んだ,古代の内部共生体に由来することを示す。長い時間の経過とともに,葉緑体の多くの特徴 ( 光合成能も ) が失われたのと同じように核も失われたと考えられる ( この例では,核変形体も残っていない ) 。
| 最初に戻る |
| メニューのページへ戻る |
February 06, 2020