生殖細胞の形成
| このページの内容 |
精子および精液
精子 spermatozoon, pl. spermatozoa は 精巣?で生産される雄の生殖細胞で,
- 受精能
- 運動能
を備えている。
これらの機能を効率よく発現できるように,精子は精密な構造をもつ。
1. 精子の形成
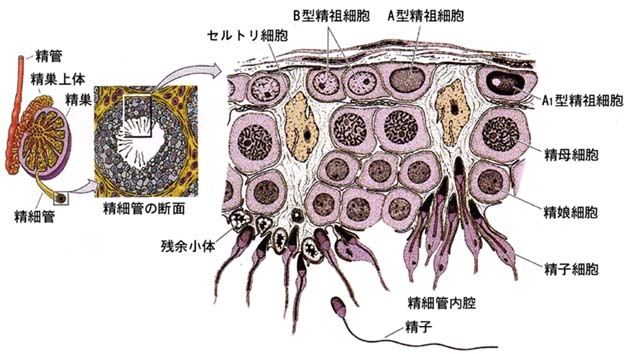 精子形成?spermatogenesis と呼ばれる一連の変化は,
精子形成?spermatogenesis と呼ばれる一連の変化は,
- 精祖細胞 spermatogonium
- 精母細胞 primary spermatocyte
- 精娘細胞 secondary spermatocyte
- 精子細胞 spermatid
- 精巣精子 testicular spermatozoon
の順に形成される変化である。
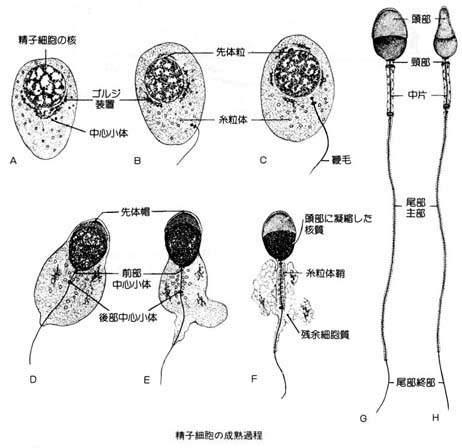
- 精祖細胞から精子細胞が形成される過程は細胞分裂によるので,精子発生?spermatocytogenesis ( 右図 ) という。
- 精子細胞から精巣精子になる過程は細胞分裂なしに精子に変態するので,精子完成?spermiogenesis (右下図)という。
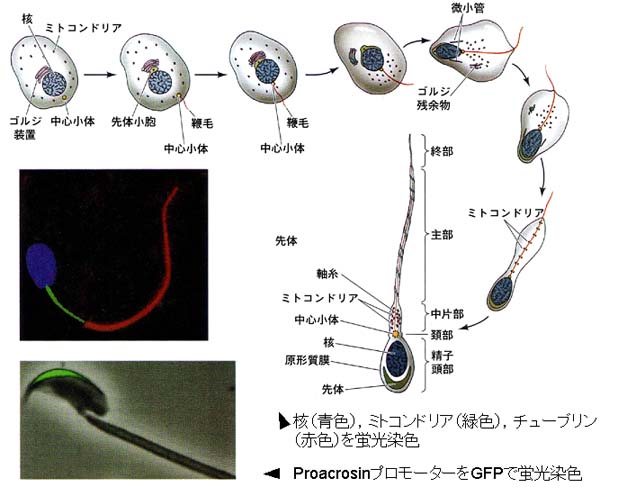
2. 精子の形態
精子は
- 頭部 head
- 頚部 neck
- 尾部 tail
- 中片部?middle piece
- 主部?principal piece
- 終部?end piece
からなる。
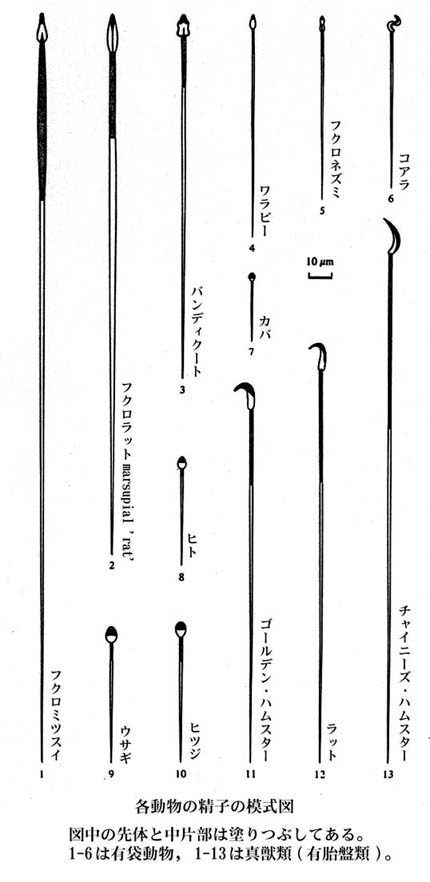
精子の形態や大きさは動物種によってかなり異なるが,ウシ,ウマ,ブタ,ヒツジ,ヤギなどの家畜ではいずれも頭部は扁平・卵円形で,全長 50 – 70?mm である。
また,ニワトリ精子の頭部は,わずかに湾曲した棒状を示す。
| いろいろな動物の精子の形態はこちら |
a. 頭部
頭部の前半は帽子状の 先体 acrosome で,後半は 後核帽 postnuclear cup で覆われている。
 先体は精子完成過程中にゴルジ装置より形成されたもので,先体内部には受精の際に卵丘細胞の結合を緩める作用をもつ?ヒアルロニダーゼ?や透明帯通過に必要なアクロシン?などの多種類の酵素を含む。
先体は精子完成過程中にゴルジ装置より形成されたもので,先体内部には受精の際に卵丘細胞の結合を緩める作用をもつ?ヒアルロニダーゼ?や透明帯通過に必要なアクロシン?などの多種類の酵素を含む。
受精に際し,精子細胞膜と先体外膜との部分的融合によって細胞膜が多孔状になる変化が見られ,内部の酵素が浸出し易くなる。この変化を?先体反応?acrosome reaction という。
b. 頚部
頚部は頭部と尾部の連結部で主として?中心小体?から生じた部分である。中片部とともに精子の運動起点をなしている。
c. 尾部
尾部は運動器官で全長を軸繊維束が貫いているが,その構成と軸繊維束をとりまく構造物の差によって,以下のように区分される。
- 中片部の軸繊維束中心微小管 2 本,周辺微小管 9 本,周辺繊維 9 本の計 20 本からなり,その外側をミトコンドリアがラセン状に取り巻いて?ミトコンドリア鞘?mitochondrial sheath を形成している。
- 主部では軸繊維束の周辺繊維が先に行くにしたがい次第に細くなって消失し,11 本の微小管のみとなる。またミトコンドリア鞘はなく,そのかわり表面は尾鞘で覆われる。
- 終部は尾部の末端の短い部分で,尾鞘がなく,微小管が直接細胞膜で包まれている。
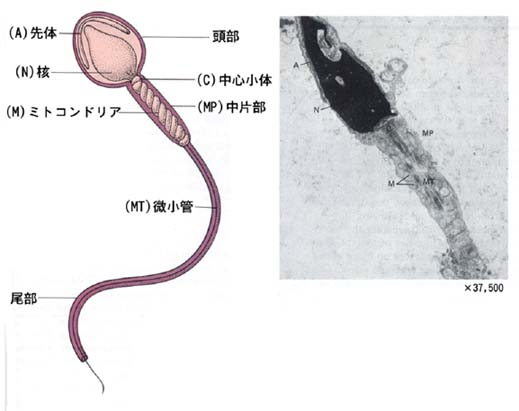 尾部の構造物のうち軸繊維束の周辺繊維は収縮して尾部を運動させ,ミトコンドリア鞘はそのエネルギーを獲得する役割を果たしていると考えられている。
尾部の構造物のうち軸繊維束の周辺繊維は収縮して尾部を運動させ,ミトコンドリア鞘はそのエネルギーを獲得する役割を果たしていると考えられている。
精巣および精巣上体から取り出した精子には,細胞質の残留物である小滴が付着しており,これを?細胞質小滴?という。
付着部位は頚部から中片部の末端までの範囲で,未熟精子の指標とされる。
3. 精子の生理
a. 運動性
哺乳動物の精子の運動は,尾部を縦走する 20 本の?微小管?の作動による。
これらの繊維束の主成分である収縮性タンパク質は,筋肉のタンパク質と化学的性質も収縮の様式もよく似ている。
射精直後の精液では,精子の運動は極めて旺盛なため,全体として渦のような流れが認められる。個々の精子の運動を見れば,長い尾部を激しく波状に動かし,頭部を回転させながら前進する。
実験的に測定した運動速度は,
- ウシで毎秒 94 – 123?mm,
- ウマで 87?mm,
- ヒツジで 50 – 80?mm,
- ニワトリで 17?mm
と報告されている。
b. 趨向性
精子運動には種々の趨向性が観察されている。
- 向流性?rheotaxis
精子に対して外部から流れを与えれば,精子の大部分はその流れに逆らって泳ぎ始める性質。 - 向触性?thigmotaxis
気泡や細胞片などの固形物が存在している場合には,精子はこれらに集まってその頭部を付着させる傾向を示す性質。 - 向化性?chemotaxis
精子には特定の化学的成分に向かって泳ぐ性質。
c. 生存性
- 雄の生殖道内における精子の生存性精巣内で形成された精子が射出するまでにはかなりの日数がかかるが,精子はこの間に徐々に成熟して運動性を獲得するようになる。精子が精巣上体内を通過して貯蔵場所である精巣上体尾部に到達するのまでの時間は,
- ラットで 15 日,
- ウサギで 4 – 7 日,
- モルモットで 14 – 18 日,
- ヒツジで 9 – 14 日であるが,個体によっても異なる。
一方,雄の生殖道内における精子の生存時間は,精子の滞留部位によって異なり,
- 精巣上体頭部では最も早く死滅し,
- 精管がこれに次ぎ,
- 精巣上体尾部でもっとも長く生存する。
- ラットで 30 – 45 日,
- モルモットで 50 – 59 日,
- ウサギ,ウシで約 60 日生存することが知られている。
- 射出精子の生存性射出された精子の置かれた環境によって著しく異なる。射出直後の精子は,活力の高い精子ほど生存能力が高く,また精子の代謝が盛んであるほど生存性,受精能力が高い。しかし,精子を長く保存するためには,精子の代謝を抑制する環境におく必要がある。現在,精液は凍結保存によって半永久的に保存可能である。
- 雌の生殖道内における精子の生存性この生存時間は動物種によってかなり違うが,一般に上記 1, 2 の生存時間に比べて著しく短い。部位別にみると,膣内で最も生存時間が短く,子宮頚部で最も長い。また,発情期では非発情期に比較して,精子生存時間が長い。
精子は雌の生殖器に入って,子宮から卵管上部に移送される間に重要な形態的ならびに機能的変化を受ける。この変化を?受精能獲得?capacitation と呼ぶ。
受精能獲得については,これを失わせる作用,すなわち脱受精能獲得 decapacitation と,さらに再現させ得る作用,すなわち再受精能獲得 recapacitation も確認されている。
d. 精子の運動性と生存性に影響する要因
- 温度
- 水素イオン濃度と浸透圧
- 各種イオン
- 光線とX線
- ガス体
e. 代謝能
精子は呼吸および解糖によって物質代謝を行い,運動維持に必要なエネルギーを生産している。
4. 精液 semen
精液は,
- 細胞成分である精子と
- 液状成分である?精漿?seminal plasma
からなっている。
精子は 精巣 で生産され?精巣上体?に蓄えられており,精漿は主として 副生殖腺?である 精嚢腺,前立腺,カウパー腺 の分泌液の混合したもので,精巣上体,精管?の分泌液も微量であるが含まれている。
a. 精液量および精子数
1 回に射精させる精液量および精子数は動物種により大きな差があり,また同じ動物種でも品種,個体,季節,栄養状態,射精頻度,精液採取方法によってかなり異なる。
b. 精子の生化学的組成
精子頭部の核 には遺伝情報を伝える DNA – タンパク質複合体 が存在し,先体?には脂質,糖,タンパク質の複合体とリソゾーム lysozome 酵素群が含まれている。
また,精子の中片部と尾部には主としてリン脂質,シトクロームおよび各種の脱水素酵素が含まれている。
c. 精漿の生化学的性状
精漿の中には,雄性生殖器および副生殖器からの分泌液と精子から漏出した成分が混在するため,他の体液とは著しく異なり,また動物種によっても著しい差がある。
無機質としては,K,Na,Ca の含量が高く,有機質では果糖,クエン酸,ソルビット,イノシット,グリセロリン酸コリン,エルゴチオネインなどが一般の体液内では見られない濃度で含まれる。
卵子
卵子は雌性生殖腺,すなわち卵巣で生産される生殖細胞で,雄性動物の生殖細胞である精子と受精し,新しい個体へ発生する。
1. 卵子の形成
雌の性細胞が分化し,成熟する過程を 卵子形成 oogenesis という。この過程は胎生期における卵祖細胞の分化に始まり,卵母細胞の成熟分裂を経て,排卵とそれに続く受精によって完了する。まず,
- 卵祖細胞 oogonium が有糸分裂によって増殖し,
- ついで 卵母細胞?primary oocyte へ分化して体積を増す。
- さらに,成熟分裂の第 1 分裂を経て 卵娘細胞 secondary oocyte と 第 1 極体 first polar body を形成し,
- ついで卵娘細胞は第 2 分裂によって 卵細胞 ootid と 第 2 極体 second polar body を生ずる。
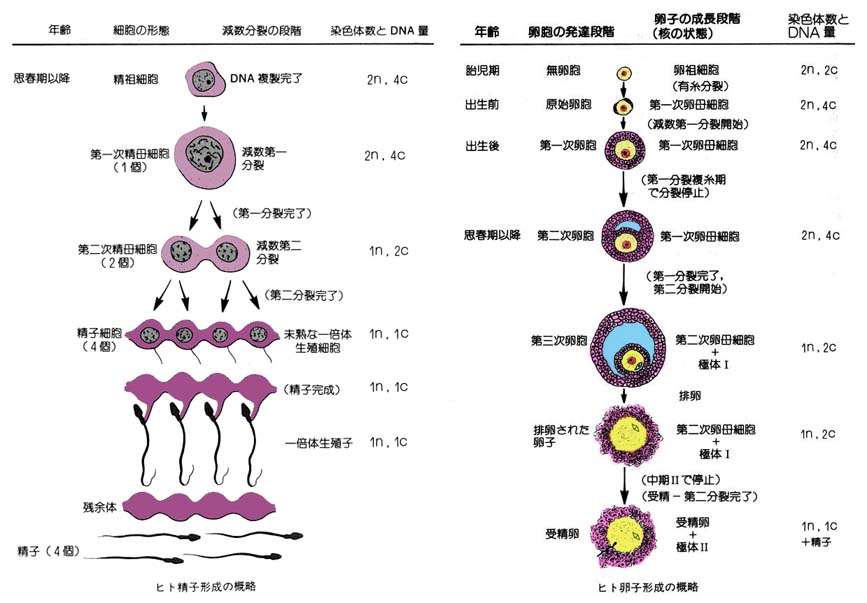
2. 卵胞発育
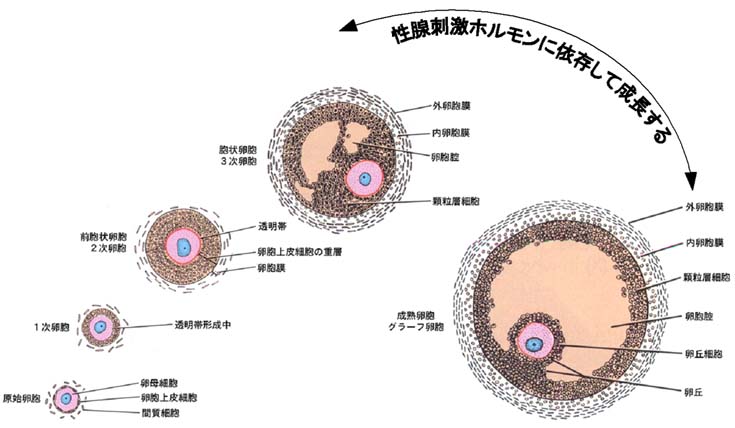
原始生殖細胞が生殖隆起への移動を完了すると,生殖細胞は直ちに扁平な細胞群に囲まれるが,この細胞群が 卵胞上皮細胞 である。卵胞の発育に伴い重層化した卵胞上皮細胞を?顆粒層細胞 と呼ぶ。
- 原始生殖細胞と,それを囲む 1 層の扁平な 卵胞上皮細胞 群からなる 1 つの単位は 原始卵胞 primordial follicle という。
- 原始卵胞内の 1 層の卵胞上皮細胞群は扁平から立方状に変わり,卵母細胞の体積も増加する。この時期を 1 次卵胞 primary follicle という。
- その後は次第に増殖分裂して重層化し,その形態も立方状あるいは円柱状となる。この時期の卵胞は 2 次卵胞 secondary follicle という。透明帯 zona pellucida はこの時期に形成される。
- 顆粒層細胞の分裂がさらに進むと卵胞内に空隙が生じ卵胞液が貯留する。卵胞腔が生じた卵胞を 胞状卵胞 と呼ぶ。
- さらに卵母細胞を取り囲んだ卵胞上皮細胞の分裂増殖によって,卵胞腔内に突出するようになる。この様な状態を 卵丘 cumulus oophorus と呼び,この部分を構成する卵胞上皮細胞を 卵丘細胞 cumulus cell という。
卵丘細胞のうち卵母細胞の透明帯と接して,卵子を直接取り囲んでいる最内層の細胞群は,放射状に配列していることから,放射冠 corona radiata と呼ぶ。 - 大きな腔を有する十分に発達した大卵胞は,とくに グラーフ卵胞 Graafian follicle といわれている。グラーフ卵胞内の卵母細胞は?卵核胞 germinal vesicle とよばれる球形の大きな核を有し,染色体は細い網目状を呈する特殊化した複糸期( diplotene 期)にとどまる。
{kind=link}
{kind=link}
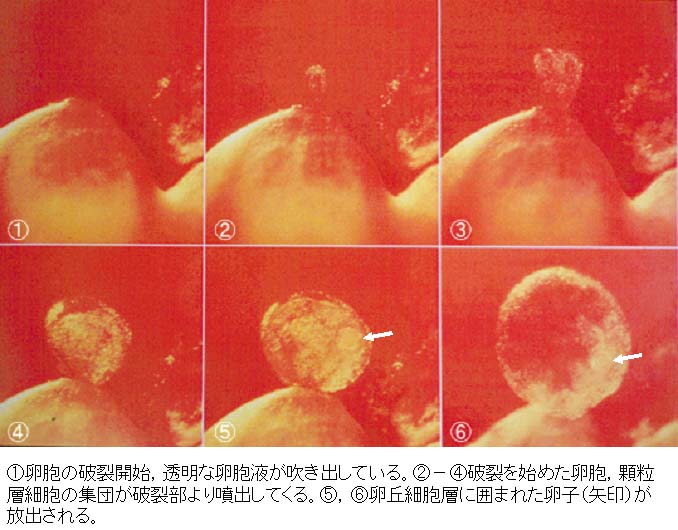
3. 排卵期における成熟分裂の再開
発情期の ゴナドトロピン( 性腺刺激ホルモン )の大量の放出によって,グラーフ卵胞の卵細胞( 卵母細胞 )の成熟分裂が再開し,発生能を有する卵娘細胞が卵胞から放出される現象を?排卵 ovulation という。
排卵は 下垂体前葉 からの ゴナドトロピン( 性腺刺激ホルモン ) の急激な放出( LH サージ )によって誘起される。
LH サージ から排卵までの時間は動物種によってほぼ一致している。この間に卵巣の内分泌的変化,卵胞壁の構造変化が同調して進行し,排卵後には黄体が形成される。
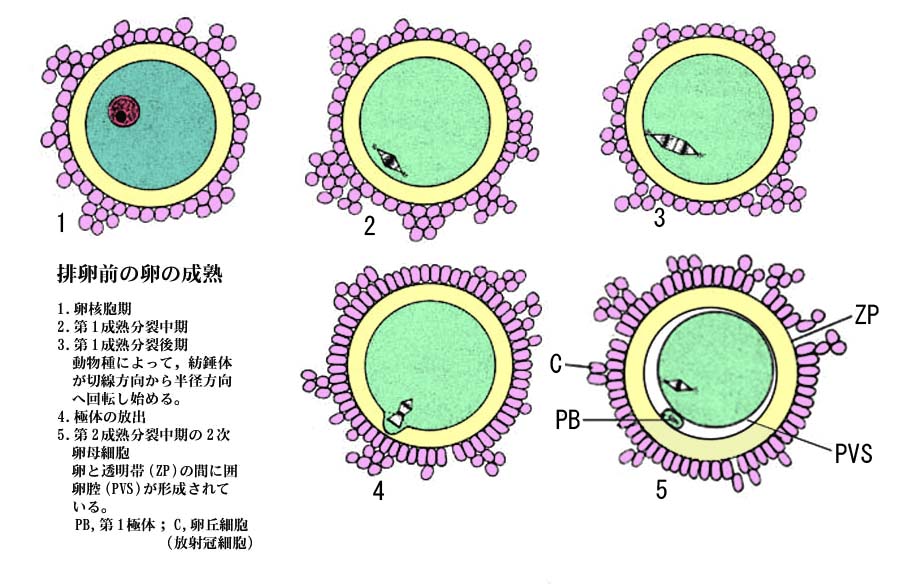
卵母細胞では長い間中断していた成熟分裂が再開する。
成熟分裂が再開すると,
- 卵核胞の核膜が壊れ,核小体も消失して,周囲の細胞質と区別がつかなくなる。 この変化を 卵核胞崩壊 germinal vesicle breakdown, GVBD と呼ぶ。
- ついで,染色体および紡錘糸が現れ,第 1 分裂の中期から終期を経て,第 1 極体を放出し染色体は半減する。第 1 極体は?囲卵腔?perivitelline space に認められる。
この段階から卵母細胞から卵娘細胞になる。 - 卵娘細胞に残った染色体は休止核をつくらずに直ちに第 2 分裂中期に進む。
ほとんどの哺乳動物の卵子はこの段階で排卵される。
成熟分裂はここで再び中断され,精子の侵入後に後期,終期という第 2 成熟分裂を終える。

| 卵の成熟過程を示す模式図は?こちら |
| ウサギの排卵の過程は?こちら |
{kind=link}
{kind=link}
4. 卵子成熟の制御機構
- 成熟/M期促進因子(Maturation/M-phase promoting factor:MPF)
○MPF は酵素活性をもつp34cdc2と制御因子サイクリンB ( cyclin B ) との複合体である。
○サイクリン B の分解が抑制されると MPF 活性が高く分裂期が維持され,逆にサイクリン B が分解されると MPF 活性が低下し,染色体は脱凝縮して核膜が形成される。
○MPF はタンパク質リン酸化酵素作用をもつ。
○活性化された MPF が種々のタンパク質をリン酸化することにより,染色体の凝縮や核膜の消失などの変化(すなわち,細胞周期の G2/M 移行)が起こる。 - MAP キナーゼ mitogen-activated protein kinase ( MAP とは分裂促進因子)
○排卵卵子では MAP キナーゼ活性が高く保たれている。
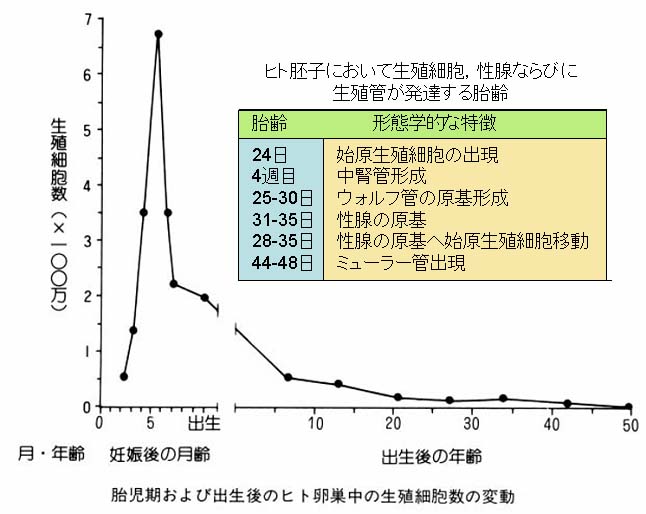
5. 卵子形成の効率
1 発情期に排卵される卵子の数は,動物種により決まっている。
単胎動物では原則として 1 個であるので,生涯に排卵される卵子数は極めて少ない。
たとえば, 1 頭の雌ウシが 15 年間に亘り休みなく 3 週間ごとに排卵を繰り返したとしても,総排卵数は約 260 個にすぎない。
出生直後の卵巣にはおよそ 6 – 10 万の卵母細胞が存在するといわれるので,排卵されるものはその僅か 0.2 – 0.4% に過ぎない。
| 女性の生殖細胞数の変動を示したグラフはこちら |
{kind=link}
| 最初に戻る |
| メニューのページへ戻る |
February 06, 2020