| このページの内容 |
核
核は真核生物にとって最も顕著な特徴を示すものである。すなわち,”真核生物” という用語は正に “真の” 核をもっていることを意味する。
核膜
核は二重の膜構造 ( 核膜 ) によって他の原形質成分 ( 細胞質 ) から隔離されているが,細胞質への通路として核膜には多数の小孔( 核膜孔 )があいている。ここでは,核膜孔複合体 nuclear pore complexes ( NPCs ) が核に出入りする分子の通過を制御している。また,核膜の内膜と外膜の間にある腔隙は小胞体系の内腔を通ってやがて細胞外液につながっている。
クロマチン ( 染色質 )
核は細胞の 染色体( 遺伝情報担体 )を含む。各染色体は一本鎖 DNA と量的にほぼ同量のタンパク質が結合したものである。集合的に,タンパク質が結合した核の DNA をクロマチン chromatin (染色質)と呼んでいる。
これらのタンパク質は,ヒストン histone と呼ばれる 5 種類の分子種 ( H1,H2A,H2B,H3,H4 ) で,正に荷電した アルギニン と リジン残基が多い塩基性タンパク質である。( アルギニンとリジンはどちらも R 基 に遊離アミノ基をもち,その陽子 ( H+ ) のために正に荷電している。)そのため, DNA の負に荷電した リン酸基 に強力に結合する。
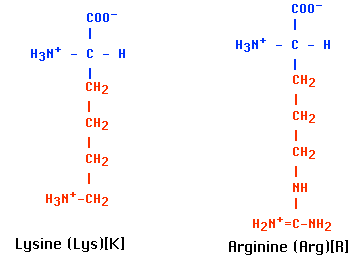{kind=link}
{kind=link}
クロマチンは極めて少量ではあるが,多種多様の 非ヒストン蛋白質 も含む。これらのほとんどは 転写調節因子 ( たとえば,ステロイドホルモン受容体 ) で,これらの DNA への結合はほんの一時的なものである。
ヌクレオソーム Nucleosome
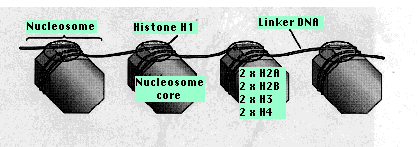 4 種類のヒストンがそれぞれ 2 分子ずつ,
4 種類のヒストンがそれぞれ 2 分子ずつ,
- H2A
- H2B
- H3
- H4
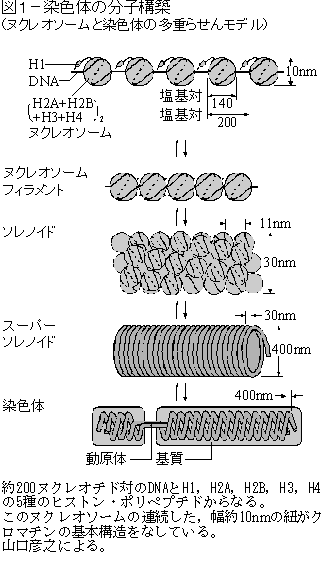
すなわち,ヒストン 8 分子がタンパク質のコア,すなわち ヌクレオソーム nucleosome コアを形成する。この周りに 147 塩基対の DNA が 2 巻きしている。
10-60 塩基対の DNA がヌクレオソーム間を連結している。その連結部分には ヒストン 1 ( H1 )が 1 分子結合する。
DNA へのヒストンの結合は DNA の特別なヌクレオチド配列によるのではなく,ヒストンのアミノ酸配列が決定的に働いている。ヒストンは進化の過程で最もよく保存された分子の例である。子ウシのヒストン 4 ( H4 )はエンドウ豆の H4 と 102 個のアミノ酸鎖のうち 2 個異なるだけである。
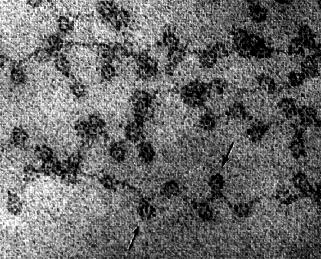
右の電子顕微鏡写真はニワトリの赤血球の核のクロマチンを示す ( 哺乳動物と違って,鳥類は成熟した赤血球に核をもっている ) 。矢印はヌクレオソームを示す。ヌクレオソームが糸に通したビーズのように配列している。
DNA を核に納めるためにはヌクレオソームを形成しただけではまだ不十分である。ヒトの 46 本の DNA 分子 ( 合計で 1 m を超える長さとなる ) を直径 10 µm 程度の核に納めるためには,さらにぎっしり折り込む必要がある。
- ヒストン・コアの露出した “尾部” 間の相互作用によって,ヌクレオソームが直径 30 nm のぎっしり折り込んだ繊維となる。
- これらの繊維はさらに複雑な高次構造に折り込まれるが( 仮想的な構造 が提案されている ),細胞分裂に入って染色体が凝縮し,太さを増す機構はまだ十分に解明されていない。また,遺伝子の活性に伴い部分的に構造が変化する仕組みも分かっていない。
{kind=link}
ヒストンの修飾
ヒストンのアミノ酸配列が変化しないが,化学修飾のために個々のヒストン分子の構造が変化することがある。
これらには,以下の修飾がある:
{kind=link}
{kind=link}
{kind=link}
ヒストン分子の 75 – 80% がコア部分に組み込まれているが,残りは N-末端 に “尾部” としてコアからぶら下がっている( 図には示していない )。
化学修飾は,とくに H3 と H4 の “尾部” に起こり,その変化は可逆的である。
たとえば,アセチル基が,
- ヒストン・アセチルトランスフェラーゼ histone acetyltransferase ( HAT ) と呼ばれる酵素によって 追加 される。 また,
- ヒストン・脱アセチル化酵素 histone deacetylases ( HDAC ) によって 除去 される。
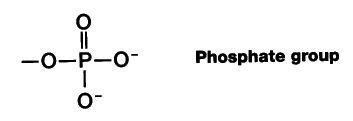
たいていヒストン尾部のアセチル化は遺伝子 転写 が活性化するクロマチン領域に起こる。 強い負の荷電をもつ DNA と強い正の荷電をもつヒストン間の結合力を低下させるために,リジンの正の荷電をアセチル化することによって中和するという仕組みのようである。
まとめ
- メチル化によって,リジン ( またアルギニン ) の荷電を中和し,その領域の遺伝子転写を促進または抑制することが可能である。
- H3 の第 4 アミノ酸リジンのメチル化は遺伝子の 活性化 に関連がある。その上,
- H3 の第 9 アミノ酸リジンのメチル化は遺伝子の 不活化 に関連する。
- H3 と H4 のアセチル化は遺伝子転写を低下させることがある。
- リン酸基が付加されると,染色体がさらに凝縮する( 有糸分裂や減数分裂時のように )。A
いずれにせよ,ヒストンはクロマチンの活動的な構成分であって,単なる DNA の梱包物質ではない。
真正クロマチンとヘテロ ( 異質 ) クロマチン
分裂間期 では,クロマチンの構造は ( 光学顕微鏡レベルで ) ほとんど見えない ( ショウジョウバエや他のハエの多糸染色体などの特殊な例を除いて ) 。しかし,核におけるクロマチンの密度が異なる。詰め込まれている程度差によると思われる。すなわち,
- 密度の高い部分をヘテロクロマチンと呼ぶ。
- 密度の低い部分を真正クロマチンと呼ぶ。
ヘテロクロマチン は,
- 以下のような,遺伝子がほとんど,または全く存在しない染色体領域に認められる。
- 動原体 centromere
- テロメア telomere
- トランスポゾンや他の “junk” DNA
- 詰め込み密度が高い。
- 細胞周期 S 期 の後期に複製される。
- 減数分裂 における乗り換えが少ない。
- ヘテロクロマチンにある遺伝子は一般的に不活化されている。すなわち,転写 されない。そして,以下の状態を示す:
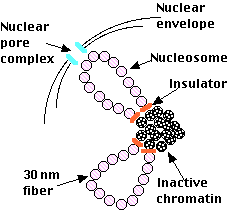 真正クロマチン は,
真正クロマチン は,
- 遺伝子転写が活発な染色体領域にみられる。
- 30-nm 繊維 が緩やかにループ状に折り畳まれている。
- インシュレーター insulator によって隣接する ヘテロ クロマチンと分離されている。.
インシュレーターについてはこちら - このループがしばしば 核膜孔複合体 の付近に認められる ( 遺伝子転写産物が細胞質へ達しやすくなっているようだ )。
右図は脊椎動物細胞の 間期 における真正クロマチンをヘテロクロマチンの様子を示した仮想的なモデルである。
ヌクレオソームと転写
プロモーターがヌクレオソームでふさがれると,転写調節因子がその プロモーター に結合できない。少なくともある遺伝子にとって,転写調節因子 を組み立てる最初の機能の 1 つが,プロモーター領域の DNA 分子に組み込まれているヌクレオソームをスライドさせることである。こうすることによって,転写調節因子がプロモーターに結合できる。
核小体 Nucleolus
分裂間期には染色体が拡張した状態にあり,染色体 1 本以上 ( ヒトの細胞では 10 本 ) が核小体と呼ばれる球状の集合体へループを伸ばす。ここでは,リボソームの大・小のサブユニットの組み立てに使用される ( 4 種類のうちの ) 3 種類の RNA 分子 ( 28S,18S,5.8S )が合成される ( 5S rRNA 分子は核内の他の部位で合成される ) 。
10 本の染色体上 ( ヒト ) にある数百から数千の直列に並んだリボソーム遺伝子 ( rDNA ) から ( RNA ポリメラーゼ I によって ) 28S,18S,および 5.8S リボソーム RNA が転写される。これらの 10 本の染色体の rDNA 領域が核小体に集まる。
これらが合成されると,多量の各種リボソームタンパク質 ( リボソームの大・小サブユニットの組み立てに使われる ) に rRNA 分子が結合する。
しかし,すべてのタンパク質はリボソームによって細胞質で合成される。したがって,核内で形成されたリボソームの部品を細胞質へ輸送する機構が存在するはずである。核膜孔複合体の機能の一つがこれに当たる。
核膜孔複合体 Nuclear Pore Complexe ( NPC )
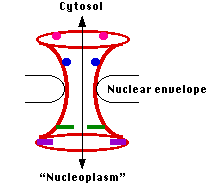
核膜には多数の核膜孔がある。
それぞれが ヌクレオポリン nucleoporin と呼ばれる種類の異なるタンパク質 ( 酵母で 30 種類,おそらく脊椎動物では 50 程度 ) から構成されている。
これらが完全に組み立てられると, 核の内部にある原形質 ( “核質” nucleoplasm ) につながる水溶性チャンネルを形成する。物質が核膜孔を通過する時,これが 25 nm 程度に開口する。リボソームのサブユニットのような大きな部品が通過するには十分の大きさである。
核膜孔複合体を通過する輸送は 能動 輸送である。したがって,
- エネルギー
- 特定物質の輸送に適した各種の輸送分子
- 核膜孔複合体における結合分子 ( 図には小さな直方体と円形で示してある )
核内への輸送
すべてのタンパク質は細胞質で合成され,核が必要とするタンパク質は核膜孔複合体から核内へ輸送しなければならない。おそらく,これらのタンパク質の各々はアミノ酸の特有の配列を持っており,これが核内に入る標識となる。
核内へ輸送されるタンパク質は:
- ヌクレオソームをつくるのに必要なすべての ヒストン
- リボソームの組み立てに必要なすべての リボソームタンパク質
- 遺伝子のスイッチを入れたり切ったりするすべての 転写調節因子 ( たとえば,ステロイドホルモン受容体 )
- pre-mRNA から 機能的 mRNA へ処理するためのすべての スプライシング因子 ( イントロン領域を取り除き,エクソン領域を連結する )
核外への輸送
核から持ち出される分子ならびに高分子の構成要素は:
- > rRNA とタンパク質の両方を含むリボソーム・サブユニットメッセンジャー RNA ( mRNA ) 分子 ( タンパク質も含まれる )
- 転移 RNA ( tRNA ) 分子 ( 同様に,タンパク質も含まれる )
- 転写調節因子 ( 利用後,細胞質に戻り,再利用時に核内に輸送される )
RNA とタンパク質分子の両方が特徴的な 核外輸送シグナル をもち,これにより正しい輸送分子と結合でき,細胞質へ輸送される。
“核質”
この用語は,核の内容物を指す。たとえば,複製 と 転写 が核内の別々のところで行われていることを示す証拠がある。現在のところ,核内に存在すると思われる構造上の複雑さに関する情報は少ない。
April 2, 2020