真核生物における遺伝子制御
![]()
5 April 2016 版を翻訳一部改変
| このページの内容 |
ヒト・ゲノム・プロジェクトにおける最新の推定では,真核細胞であるヒト細胞は約 21,000 個の遺伝子をもっている。
- ある遺伝子は常にすべての細胞で発現している。これらはハウスキーピング遺伝子と呼ばれ,すべての細胞の生命維持に必要な代謝機能 ( たとえば呼吸など ) を制御するのに必要である。
- ある遺伝子は,細胞が分化の特定の経路に入る時に発現する。
- ある遺伝子は,特定の方法で分化した細胞においてのみ常時発現する。たとえば,形質細胞は抗体の遺伝子に常にそれを産生するように働いている。
- ある遺伝子は,細胞の周囲あるいは細胞内の状態が変化した時に発現する。たとえば,ホルモンが作用してその細胞の特定の遺伝子の活動を始めさせたり,終わらせたりしている。
遺伝子の発現はどのように制御されているのか?
真核生物では,DNAは非常に複雑なクロマチン構造を形成しているため,RNAポリメラーゼが単体で結合することが難しい(原核生物ではRNA単体で転写が開始される:詳細についてはこちら)。
真核生物ではいくつかの方法が知られている:
- 遺伝子の 転写 の速度 が変化する。これは最も重要で,広く用いられている戦略である。
- しかし,真核細胞は転写制御にいくつかの他の方法をさらに付け加えている。
- RNA 転写産物が核内にあるうちに処理速度を変化させている。[ RNA の作用についてはこちら ]
- mRNA 分子の安定性を変化させている。すなわち,分解される時の速度を変化させている。[ RNA 干渉についてはこちら ]
- リボソームが mRNA を ポリペプチド に翻訳する効率を変化させている。 [ 翻訳制御の例はこちら ]
タンパク質をコード化している遺伝子には,以下の部位が含まれる。
-
-
- ポリペプチドをコード化している領域のエクソン
- イントロン これは介在部分で,翻訳前に除去される。[ RNA の作用についてはこちら ];
- 転写開始部位
- 転写開始位置上流のプロモーター
- コアプロモーター (core promoter ) 転写開始部位の約 40塩基対(bp: base pairs)上流に位置する。
- 上流プロモーター (upstream promoter)もう一つは,200 bp ほど上流に位置する。
- 調節領域であるエンハンサー
- 調節領域であるサイレンサー
-
隣接した遺伝子 ( RNA やタンパク質をコード化している ) はたいてい インスレーター によって分離されている。この部位は他のプロモーターやエンハンサー ( あるいはまたサイレンサー ) のそれぞれの混乱を避ける働きがある。
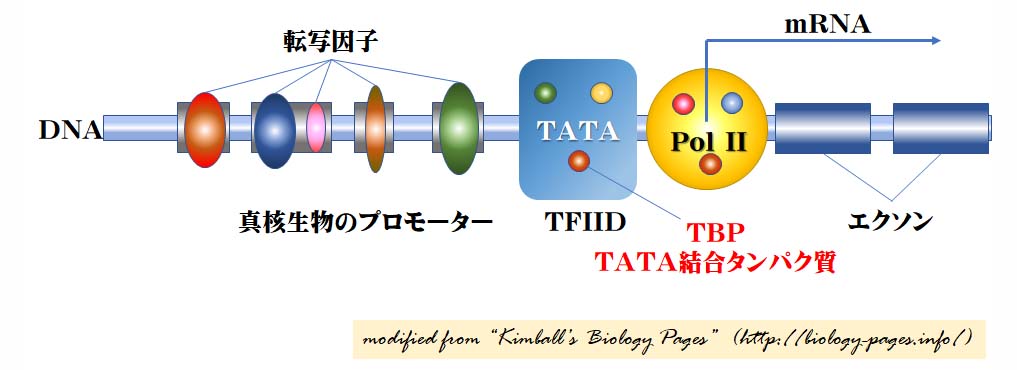
真核生物のプロモーター模式図
転写開始部位 Transcription start site
これは RNA ポリメラーゼ II ( pol II ) 分子が結合する部位である。 Pol II は 12 個の異なるタンパク質の複合体である (上図の黄色の部分に小さな円形で重ねて示してある ) 。
転写開始点から遺伝子が mRNA へ 転写を始めることになる。
コアプロモーター Core promoter
すべての真核生物の遺伝子はコアプロモーターをもつ。コアプロモーターは TATA box と呼ばれる 一連の塩基 ( TATAAAAAA ) の配列を含む。この部位には,別の 50 個のタンパク質から成る巨大な複合体が結合し,以下が含まれる。
- 転写調節因子 IID (Transcription Factor IID, TFIID) これは以下の複合体である。
- TATA box を認識して結合する TATA-結合タンパク質 TATA-binding protein ( TBP ) と,
- TBPと一部DNAに結合する他の13個のタンパク質因子
- 転写調節因子 IIB (Transcription Factor IIB, TFIIB) これはDNAとpol IIの両者に結合する。
コアプロモーターは,その構造や結合因子の種類にあまり変化は見られず,すべてのタンパク質をコードする遺伝子に認められる。これは,その構造と関連する結合因子が遺伝子によって異なる上流プロモーターと著しい対象を示す。
多数の異なる遺伝子と多種類の細胞型が同じ転写調節因子を共用している。コアプロモーター に結合するものばかりでなく,上流プロモーター に結合するものも含めてである。 特定の細胞の特定の遺伝子のスイッチを入れるものは,おそらくプロモーター部位といくつかの転写調節因子の独自の組み合わせによって決まっているのだろう。
ユニークな類似点
銀行に設置されている貸金庫とユニークな類似性がある。
貸金庫室で特定のボックスを開ける時には,2つの鍵が必要となる。すなわち,
●あなたに割り当てられたボックス用の「あなたの鍵」で,これは上流プロモーターに相当するだろう。ただし,銀行員が管理している鍵がないとボックスの解錠はできない。
●銀行員の鍵はすべてのボックスが解錠できるような仕組みを作動させることができる鍵だが,これだけでは個別のボックスは解錠できない。これはコアプロモーターに相当するだろう。
ホルモンは転写調節因子を形成することによってその効果を発揮している
ホルモンとその受容体は複合体を作り,転写調節因子の役割をもつ。
ホルモンの リスポンス・エレメント ( 反応要素 ) “response element” が結合するのがプロモーター部位なのである。[ ホルモン受容体についてはこちら ]
エンハンサー enhancer
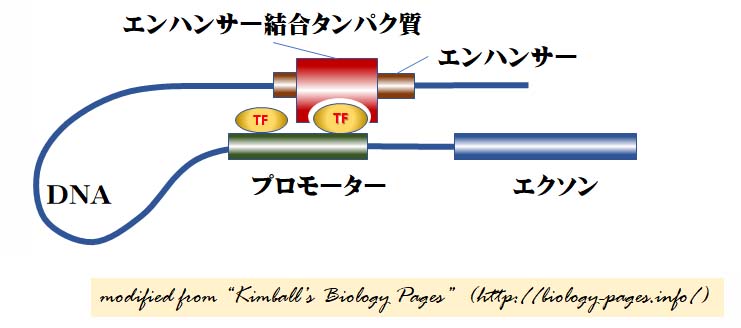
エンハンサー結合タンパク質が働きDNAループが形成される
ある転写調節因子 ( エンハンサー結合タンパク質 ) はそれらが制御する遺伝子から数千塩基も離れた DNA 領域に結合する。この結合によって,遺伝子の転写速度が増加する。
エンハンサーは上流,下流さらにはそれらが制御する遺伝子内にあることもある。
ゲノムには何千ものエンハンサーが存在するが,細胞の種類や受けた信号によって作動するものがある。
少なくともショウジョウバエでのほとんどの遺伝子は,2~3個のエンハンサーによって調節されているが,ある遺伝子では8個またはそれ以上のエンハンサーが関与している場合がある。
複数のエンハンサーはハウスキーピング遺伝子の特徴である。
エンハンサーへのタンパク質の結合が数千塩基も離れた遺伝子の転写をどのように制御しているのであろうか?
1 つの可能性は,エンハンサー結合タンパク質が,DNA結合部位と別に,遺伝子のプロモータに集まった転写調節因子に対する結合部位をもっているということである。
このことを示すために,図にはDNAをループ状に示している。
このループは以下のタンパク質によって安定化されている。
- 転写抑制因子CTCF(別名CCCTC結合因子=この塩基配列に結合するため)と呼ばれるタンパク質で,DNAの一端のCTCFは,2つの領域を結びつけているDNAの他端のCTCFと二量体を形成する。CTCFは11個のジンク・フィンガーをもつ。
- コヘーシン cohesin これは有糸分裂と減数分裂の時に姉妹染色体を固定するタンパク質複合体である。
物的証拠
マイケル・ボッチャン Michael R. Botchan 博士らは(この電顕写真を提供して頂いた)エンハンサーのこの作用モデルの物的証拠を得ている。
彼らは以下の特徴を持つ人工DNA分子を作製した。その特徴は:
- Sp1のプロモーター領域4個 一端から約300塩基。Sp1は塩基配列 5’-GGGCGG-3’ (これは多くの遺伝子,とくにハウスキーピング遺伝子のプロモーターに見つかっている塩基配列である)に結合するジンク・フィンガー転写因子である。
- 他端から800塩基の5個のエンハンサー領域をもつ。これらにはE2と呼ばれるエンハンサー結合タンパク質が結合する。
- 両者間の1860塩基対のDNA
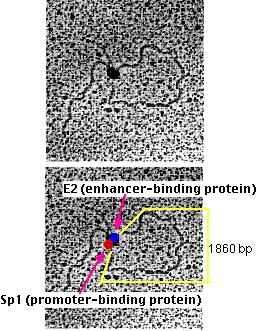
Botchan 博士らの人工DNA分子の実験結果を示す透過型電子顕微鏡像
これらのDNA分子をSp1とE2の混合液に加えると、DNAは引き込まれおよそ300塩基と800塩基の「尾」をもつループを形成することを電子顕微鏡分析により明らかにした。
各々のループのあとに2個の球状物が認められた。一つはSp1(赤色)でもう一つはE2(青色)分子である。(2枚の写真は同一のもので,下の写真には説明のためのラベルを付してある。)
プロモーター領域やエンハンサー領域を欠如した人工DNA分子,あるいは突然変異型の領域をもつDNA分子は,2種類のタンパク質と混合してもループを形成することはなかった。
ループ形成の意義
プロモーターの近くにエンハンサーを持ってくる(あるいは,プロモーターを他のプロモーターの近くにもってくる)染色体のループ形成は,一緒に実行しなければならない遺伝子グループの発現(または抑制)を確実にするメカニズムのようである。
ホルモンのようなシグナルの到着に反応して,何百もの異なる遺伝子をオンにする(またはオフにする)ことが起こり,その結果産生された遺伝子産物により,細胞が適切に反応することが可能となる。
このことを達成するために,適切な遺伝子座位をもっている染色体の情報を「転写工場」にダイナミックに送り込んでいるようだ。
細菌のオペロンによって同時に制御される複数の遺伝子発現に匹敵する機構を真核生物で見ていることになる。
サイレンサー silencer
サイレンサーは,エンハンサーと同様に当該の遺伝子から数千塩基も離れて位置する DNA の制御部位である。しかし,転写調節因子が結合した時,サイレンサー領域は抑制させる。
インスレーター insulator
インスレーターは,
- 42 塩基程度の DNA 配列で,
- 隣接する遺伝子または集団で遺伝子が存在する部分の
- エンハンサーとプロモーター,または
- サイレンサーとプロモーターの間に位置する。
これらの機能は,隣接する遺伝子の活性 ( あるいは抑制 ) 化によって当該の遺伝子が影響を受けるのを防いでいる。
例:
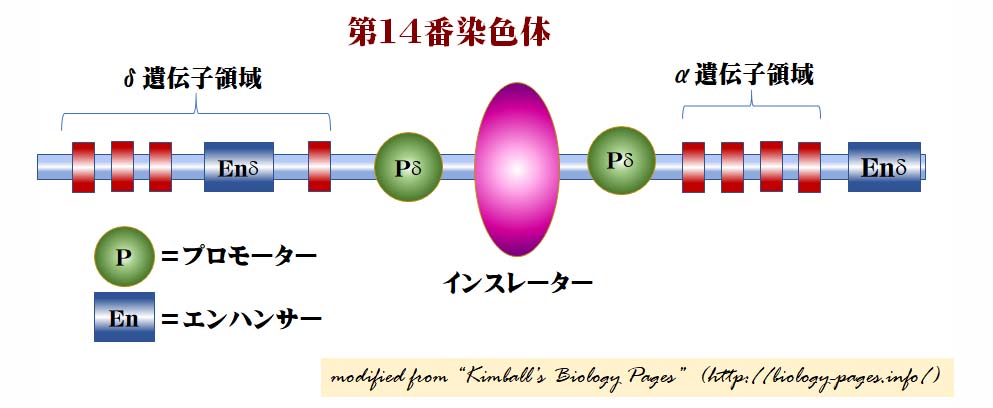
ヒト第14番染色体上にある抗体に対するT-細胞受容体(TCR)の模式図
抗体に対するgamma/delta T-細胞受容体(TCR)のデルタ鎖に対する遺伝子のプロモーターのエンハンサーは, alpha/beta T-細胞受容体のアルファ鎖のプロモーター近くに位置する ( ヒトの第14 染色体上 ) 。T細胞はどちらか一方を選ぶことになるが,alpha 遺伝子プロモーターと delta 遺伝子プロモーター間にはインスレーターが存在して,一方の活性化が他方に及ばないようになっている。
これまで見つかっている脊椎動物のすべてのインスレーターは,CTCFタンパク質が結合した時にだけ作用する。
別の例:
哺乳動物(マウス,ヒト,ブタ)では,インスリン様成長因子2(insulin-like growth factor-2,IGF2)遺伝子は父親由来の遺伝子のみが発現し,母親由来の遺伝子は発現しない。この現象は,ゲノム刷り込み (genomic imprinting) として知られている。
その仕組みは,以下の通り。
母親由来の対立遺伝子はIGF2遺伝子プロモーターとエンハンサーとの間にインスレーターをもつ。父親由来の対立遺伝子もそうなのだが,父親由来の遺伝子の場合には,インスレーターはメチル化されている。
したがって,CTCFはインシュレーターに結合できないので,エンハンサーは作動して父親由来のIGF2遺伝子のプロモーターのスイッチを入れることが可能となる。
ブタの畜産的に重要な品種の多くは,脂肪に対する骨格筋の比率を増やす遺伝子をもつように改良されてきた。
この遺伝子は配列決定されて,IGF2の対立遺伝子であることが分かっている。そして、それはそのイントロンの1つで起こった単一の点突然変異を含んでいる。
この突然変異をもつブタは、骨格筋(肝臓ではない)で、高濃度のIGF2 mRNAを産生している。
このことから,表現型に影響を及ぼすために、遺伝子のタンパク質をコードしている領域に突然変異を起こす必要はなく,遺伝子の非コード領域の突然変異でも遺伝子発現の調節過程で影響することが明らかとなった(この場合では,肝臓ではなく筋肉に変化があった)。
| ゲノム刷り込みについてはこちら |
| 前核生物における遺伝子制御についてはこちら |
| 最初に戻る |
| メニューのページへ戻る |
March 06, 2020