遺伝子発現: 転写
![]()
2 September 2013 版を翻訳一部改変
| このページの内容 |
大部分の遺伝子はそれらがコード化しているタンパク質として発現する。この過程は 2 つの流れで起こる:
- 転写 = DNA → RNA
- 翻訳 = RNA → タンパク質
遺伝情報は DNA → RNA → タンパク質 の方向にのみ流れるという生物学のセントラル・ドグマが確認されている。
概要を示す模式図
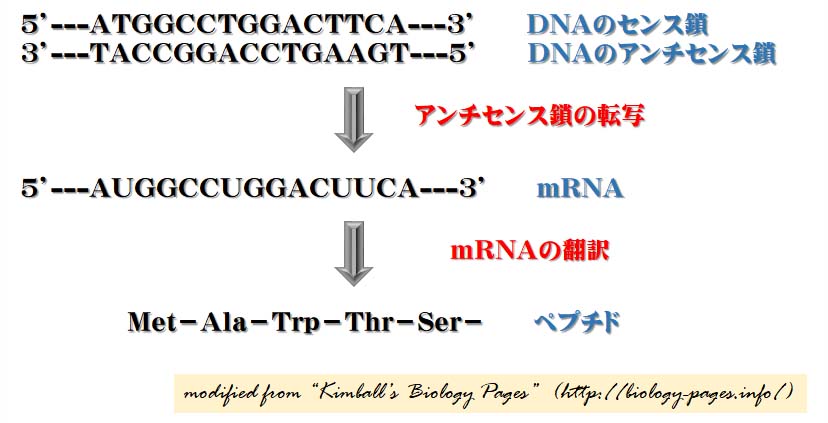
遺伝情報は DNA → RNA → タンパク質 の方向にのみ流れるという生物学のセントラル・ドグマを示す模式図
遺伝子の転写:DNA → RNA
DNA 自身の 複製 replication の時と同じように,DNA が RNA の合成のためのテンプレート ( 鋳型 ) として使われる。
転写のステップ
- 50個ほどの 転写調節因子 transcription factors が プロモーター promoter 部位( たいてい転写される遺伝子の 5′ 末端側 )に結合する。
- RNA ポリメラーゼ RNA polymerase が転写調節因子複合体に結合する。
- これらの共同作業によって,DNA 二重らせんが局所的に解離する。
- RNA ポリメラーゼがそのうちの 1 本鎖に 3′ → 5′ 方向へ移動しながら読み始める。
- 真核生物のタンパク質をコードしている遺伝子では,RNAポリメラーゼ(Pol II)の前のヌクレオソームが除去され,タンパク質の複合体がこれに関与する。DNAが転写され,Pol IIが進行したあとに,この複合体がヌクレをソームに置き換わる。
- DNA鎖に沿ってRNAポリメラーゼが進行するにつれ,リボヌクレオチド三リン酸を基質に RNA が合成される。
- 塩基の対合 のルールに従って,それぞれのリボヌクレオチドが挿入され,相補的な RNA を合成する。すなわち,DNA 鎖上の C,T,G,A の各塩基が,それぞれと相補的に対合する G,A,C,U ( ウラシル ) として RNA 鎖上に写し取られる。RNA鎖にTはない。
| 品質管理 時折、RNAポリメラーゼは誤った(塩基の対合ルールに一致していない)リボヌクレオチドを選んで、挿入する。 これがバクテリアに(そして、おそらくすべての生物において)起こるとき、この酵素は同じ動作を直ちに繰り返して、誤ったヌクレオチド(それともう一つ前のもの)を取り除いて、再びやり直す。 (Scienceの2006年7月28日号で,Zenkinらが報告している)。 |
- RNA 鎖は 5′ 末端から 3′ 末端の方向へ伸長する。
- リボヌクレオチド三リン酸が伸張中の 3′ 末端に付加され,2 個のリン酸基が除去される。
- 転写が完了した場合,RNA ポリメラーゼから転写産物が放出され,その直後にRNA ポリメラーゼがDNA鎖から遊離する。
DNA 分子のどこででも,どちらの 1 本鎖でもテンプレートとして用いられる。すなわち,ある遺伝子がこちらへ,他の遺伝子ではあちらへというふうにである(そして,希ではあるが注目に値する事例では,両方のDNA鎖上の遺伝情報を含む二重らせんの同じセグメントで同時に起こっていた)。ただし,いかなる場合でも RNA ポリメラーゼは 3′ → 5′ 方向へ移動しながら作用する。
| 2001 年 1 月 4 日号の Nature で, RNA ポリメラーゼが DNA 二重鎖に沿って移動することが報告されている。少なくとも体外培養の条件下で RNA ポリメラーゼが移動しない時は,DNA 分子を回転させることによって,DNA 分子を先に進ませるようである。体内でも同じ現象が起こるのかは分かっていない。 |
RNA の種類
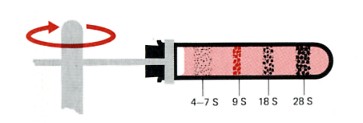
ウサギ赤血球の前駆細胞から抽出されたRNAの高速遠心分離によって生じた沈降パターン
ウサギ赤血球の前駆細胞から抽出されたRNAの高速遠心分離によって生じた沈降パターン。
別々のバンドは、RNAの特定のクラスを表わす。転移RNAバンドは約4Sの沈降係数である。
真核生物のリボソームRNAは,5S,5.8S,18Sならびに28Sの沈殿係数をもつ。(沈降係数(S)が大きいほど、分子量は大きいが、比例はしていない。)
沈降係数9SのRNAはヘモグロビン合成のためのメッセンジャーRNAで,これの細胞で合成される主要なタンパク質である。
ほとんどの種類の細胞では,メッセンジャーRNAはとても不均一で,6Sから25Sまでの範囲のRNAが少量含有されている。
真核生物の核では,いくつかの種類の RNA が合成される:
-
- メッセンジャー RNA ( mRNA )
- リボソーム RNA ( rRNA )
- 転移 RNA ( tRNA )
- 核内低分子量 RNA small nuclear RNA ( snRNA )
- 核小体内低分子量 RNA small nucleolar RNA ( snoRNA )
- ミクロRNA ( miRNA ). 非常に小さな ( 約 22 ヌクレオチド ) RNA 分子で,メッセンジャー RNA ( mRNA ) の発現を制御している。 [ 詳細についてはこちら ]
- 長鎖ノンコーディングRNA long noncoding RNA( lncRNA )
メッセンジャー RNA ( mRNA )
これが後でポリペプチドへ 翻訳 される。
mRNA はそれがコードするポリペプチドの大きさを反映して様々な大きさが作られる。
ほとんどの細胞は,何千もの異なるmRNA分子(細胞によって必要なペプチドに変換される各々)を生成する。
すべての細胞に必要なハウスキーピング “housekeeping” タンパク質 ( たとえば,解糖 に必要な酵素など ) をコードしている多くの mRNA はほとどんどの細胞で共通している。
他の mRNA は特定の細胞型に特有のもので,その細胞の機能に必要なタンパク質をコードしている ( たとえば,赤血球細胞における ヘモグロビン hemoglobin の mRNA など ) 。
リボソーム RNA ( rRNA )
mRNA が翻訳されてタンパク質が合成される場であるリボソームの構築に用いられる。
これには 4 種類ある。真核生物では,
- 18S rRNA – リボソームの 小サブユニット に含まれ,約 30 種類のタンパク質と複合体を形成している。
- 28S, 5.8S, ならびに 5S rRNA – リボソームの 大サブユニット に含まれ,約 45 種類のタンパク質と複合体を形成している。
S の数字は沈降係数 ( S はスベドベリ単位,1S = 1×10-13秒 ) で,数字が大きければ大きいほど分子が大きい ( ただし,比例はしない ) 。
28S,18Sならびに5.8S 分子は、一つの遺伝子の同一コピーから生成された単一の一次転写産物(真核生物でプロセシングされる前の mRNA)の処理によって生産される。
5S 分子は,同一の遺伝子の異なる一群から生成される。
転移 RNA ( tRNA )
伸張中のポリペプチドにアミノ酸を運搬する RNA 分子。
一般的な真核細胞では 32 種類の tRNA がある。
- それぞれは個々の遺伝子産物である。
- 小型の RNA 分子で ( 約 4S ),73 – 93 個のヌクレオチドを含む。
- お互いに対合した多くの塩基が二重らせんを形成している。
- 対合していない部分は3つのループを形成している。
- 20 種のアミノ酸の 1 つが異なった tRNA 分子と結合する( 3′ 末端側で )。ほとんどのアミノ酸は1 個から数個程度の tRNA 分子が対応する。
- ほぼ中央部分のループの3 塩基よりなる部位がアンチコドンと呼ばれる。
- アンチコドンとmRNA分子の相補コドンの間の塩基の組合せにより,成長するポリペプチド鎖に正しいアミノ酸が渡される。 [ 翻訳についてはこちら ]

酵母のアラニンtRNAの構造を示す模式図
塩基の対合によって形成されたラセン状を示すアームが4個ある。
tRNA分子(上部)にアラニンを結合させる酵素(この酵素にはATPが必要である)に左のループが結合している。
アンチコドン・ループは、メッセンジャーRNAの上で適切なコドンを認識する3つのヌクレオチドを運ぶ。
丸で囲まれた文字は,実際のヌクレオチドが示されるものの化学修飾形であることを示す。
酵母のフェニルアラニンtRNAの立体像
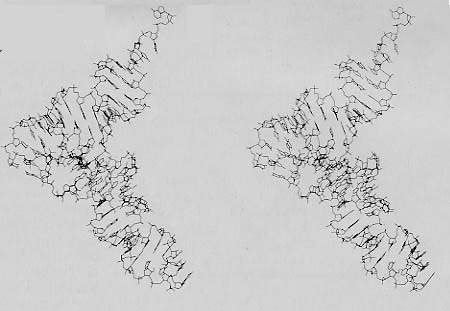
酵母のフェニルアラニンtRNAの立体像
フェニルアラニンが付着している3’端部は,右上にある。
アンチコドンは,右下にある。
2つのイメージを融合させるには、あなたの左目が左のイメージだけを,あなたの右目が右のイメージだけを見るように、2つの像の間に固い紙またはボール紙を立てると良い。
繰り返し練習すると,あなたは紙などの助けがなくても左右のイメージを融和させることができる様になる。
(Kim博士のご厚意による。出典:Sussman and Kim,Science 192:853-858, May 28, 1976.)
核内低分子量 RNA ( snRNA )
mRNA, rRNA, ならびに tRNA の遺伝子の DNA 転写によって,高分子の前駆体が形成される(一次転写産物 primary transcript)。そして,これが核内で処理され,機能的な分子に変化して細胞質へ移動する。この時の処理過程のいくつかが snRNA によって仲介される。
snRNA に対して約 12 個の異なる遺伝子が同定されており,それぞれが複数コピー存在している。
snRNA は他の種類の RNA の処理にいろいろな役割を果たしている。たとえば,いくつかの snRNA は スプライセオソーム spliceosome の一部を構成する。これは,イントロンを除きエクソンをつなぐことによって, pre-mRNA を mRNA に変換する時に働く。[ 詳細については RNA プロセッシングを参照 ]
核小体内低分子量 RNA ( snoRNA )
名前の通り,この分子量が小さな RNA ( 60~300 ヌクレオチドから成る ) は 核小体 nucleolus に認められ,以下の機能を果たしている。
- あるものは 28S, 18S, ならびに 5.8S 分子の前駆体である大型 RNA を分割するのを助け,リボソームを構築するのに働く。
- 他のものは rRNA,tRNA,ならびに snRNA 分子のヌクレオチドを化学的に修飾する ( たとえば,リボースにメチル基を付加するような ) 。
- あるものは mRNA 前駆体の選択的スプライシング alternative splicing に関与して,いろいろな成熟 mRNA を生成している。
- また,他のものは テロメア telomere の合成のためのテンプレートとして働く。
脊椎動物では,snoRNA は RNA プロセッシング の過程で除去されたイントロンから作られる。
長鎖ノンコーディングRNA(lncRNA)
メッセンジャーRNAだけがポリペプチドをコード化している。そして,他のすべての種類のRNAは、ノンコーディングRNAと呼ばれている。
rRNA、snRNA,ならびにsnoRNAに加えて,lncRNAsとして分類される200個のヌクレオチドより長い転写産物の多種類の集団があり,ヒトでは10,000個以上といわれている。
これらのほとんどの機能はまだ解明されていない。しかし、ある種のlncRNAは,以下のような多様な活動の調節に参加していることが分かってきた。
- スプライシング
- 翻訳
- 刷り込み,ならびに
- 転写
- XIST:XIST RNAは,数千のヌクレオチドからなり,雌における 2 本の X 染色体の一方を不活化する。
[ X 染色体不活化の詳細についてはこちら ] - あるlncRNAは、遺伝子のエンハンサー領域とプロモーター領域を接近させ(「ループ形成」参照),遺伝子転写を調節している。
- XIST:XIST RNAは,数千のヌクレオチドからなり,雌における 2 本の X 染色体の一方を不活化する。
これらの機能について不明の点が多いが,総合すればノンコーディングRNAは核で進行している転写の4分の3に関与しているだろう。
RNA ポリメラーゼ
RNA ポリメラーゼは巨大な,複数のサブユニットからなるタンパク質複合体である。
真核細胞では, 3 種類が知られている。
- RNA ポリメラーゼ I ( Pol I ) – これは,28S, 18S, ならびに 5.8S 分子のrRNA前駆体を合成する(RNAポリメラーゼの中で一番活動している)。
- RNA ポリメラーゼ II ( Pol II ) – また RNAP II とも呼ばれる)- タンパク質をコードしている遺伝子をmRNA にを合成する( またsnRNA も合成する)。
- RNA ポリメラーゼ III ( Pol III ) – 5S rRNA 遺伝子とすべての tRNA を合成する。
| サイエンスの2001年6月8日号の2つの注目に値する報告には,Pol IIの構造と,それがどのようにDNAのアンチセンス鎖を使ってmRNA鎖を合成しているかについて詳細が明らかにされている。 |
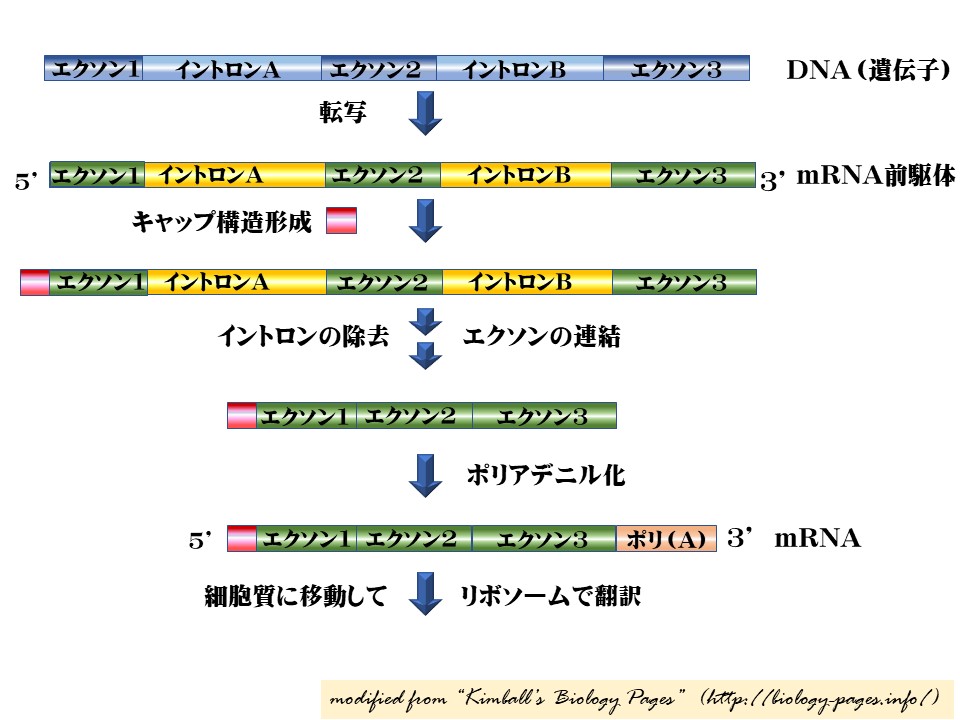
RNA プロセッシング: mRNA 前駆体 → mRNA
核において生成された一次転写産物はすべて,プロセッシングを受けないと,細胞質に送り込まれる機能的な RNA 分子にならない。ここでは,mRNA 前駆体から mRNA へのプロセッシングの各段階についてみてみよう。
RNA プロセッシング: mRNA 前駆体 → mRNA
真核生物のほとんどの遺伝子は断片化されている。
既知のタンパク質の遺伝子を読み込み枠のルールに従って解読すると,その遺伝子の実際のタンパク質産物には必要がないDNAの周期的な配列に遭遇する。
それはRNAに転写されるが、タンパク質に翻訳されない,そのようなDNAの配列はイントロンと呼ばれている。タンパク質のアミノ酸をコードしているDNA配列はエクソンと呼ばれている。
例えば:
- ニワトリで見られるあるタイプのコラーゲンの遺伝子は,52個のエクソンに分断されている。
- ジストロフィンの遺伝子(この遺伝子はX染色体に存在するので,これが変異した男子では筋ジストロフィー症が発症する)は,79個のエクソンを持っている。
- rRNAとtRNAの遺伝子でさえ,イントロンによって分断されている。
- ヒトゲノムは約180,000のエクソンを含むものと見積もられている。
- 現在,ヒトの遺伝子は21,000と推定されているが,我々の遺伝子の平均エクソン数はおよそ9個である。
一般に,イントロンはエクソンより長い傾向がある。真核生物における平均的なエクソンの長さは 140 ヌクレオチドであるが,ヒトのあるイントロンでは 480,000 個のヌクレオチドの長さにもなる。
イントロンが除去されることをスプライシングと呼び,mRNAを合成する際の必須の過程といえる。
RNAプロセシングには,キャップ構造の形成,RNAスプライシング,ポリA尾部の形成の3つの過程がある。
- キャップ構造 の合成. 転写開始後にキャップ構造と呼ばれる 7 – メチルグアノシンからなる構造が,この新生 5′ 末端に付加される。キャップ構造は RNA が 5′ 末端から分解してしまう酵素によって作用を受けるのを妨げている。
- mRNA 前駆体に存在する イントロン が段階的に除去され,残った エクソン のスプライシング(連結)。 ほとんどの真核生物では遺伝子が分断されているためにこの過程が必要となる。 mRNA 前駆体が RNA ポリメラーゼ II ( RNAP II ) によって生じる時のこの過程が起こる。
- ポリ ( A ) 尾部の合成. これは アデニン ( A ) の配列である。転写が完了した時,転写産物はある部位で切断され( 終結部位の前の数百ヌクレオチドのようである ),そして poly ( A ) 構造が剥き出しになった3’末端に付着する。これによって,mRNA が機能を持つ分子となる。そして細胞質へ移動する準備ができたわけである。( 初めにできた転写産物の残りは分解され,RNA ポリメラーゼが DNA から離れていく。)
mRNA の切断と結合は非常に正確に行われる。もし 1 個のヌクレオチドでもイントロンのが残っていれば,あるいはエクソンから取り除かれれば,その部位以降の リーディング・フレーム reading frame がシフトしてしまい,コドンが変わるためアミノ酸配列が全体的に異なるものとなってしまう ( しばしば,リーディング・フレームがシフトしたために停止コドンに変わることがあり,アミノ酸の配列が途中で中断してしまうことが多い ) 。
イントロンの除去とエクソンの結合は スプライソーム spliceosome の働きによって行われる。これは,5個の snRNA 分子と約 145 個の異なるタンパク質の複合体である。
ほとんどの mRNA 前駆体におけるイントロンは GU で始まり,AG で終了する。おそらく,この短い配列がスプライソームを誘導するものと考えられている。
物的証拠
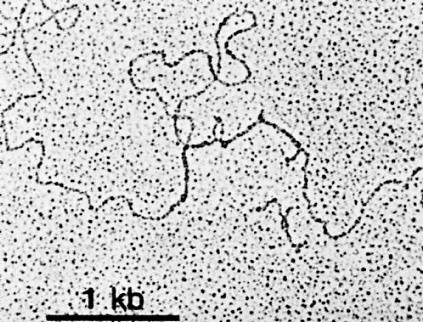
抗体分泌細胞のクローンから得たmRNAを同じ細胞の一本鎖 DNAと混合して形成された mRNA-DNA 混成分子の電顕像
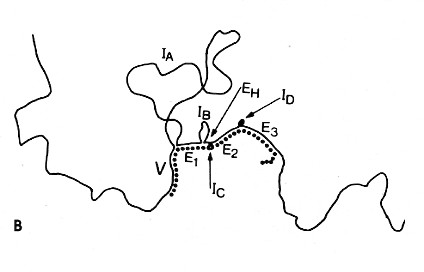
電顕写真の説明図
実線はDNAを,破線はmRNAを表す。IA,IB 等のループは,抗体の重鎖をコードするエクソンから分離したイントロンを表す。
上の写真は,抗体分泌細胞のクローンから得たメッセンジャーRNA(mRNA)を同じ細胞の一本鎖 DNAと混合して形成された mRNA-DNA 混成分子の電顕像である。バー・スケールは1,000塩基長を示す。
下の模式図は,電顕写真の説明図である。実線はDNAを,破線はmRNAを表す。IA,IB 等のループは,抗体の重鎖をコードするエクソンから分離したイントロンを表す。すなわち,
- V=V領域(可変領域)
- E1=第1C領域(CH1)ドメイン
- EH=ヒンジ領域
- E2とE3=2つのC領域(CH2とCH3)をコードするヌクレオチド
ハイブリッドを形成していない領域はポリA尾部である。
[From R. Maki et al., Proc. Natl. Acad. Sci. USA 77:2138, 1980.]
選択的スプライシング Alternative Splicing
多くのタンパク質の mRNA 前駆体の処理が,いろいろな細胞のいろいろな経路で,あるいはまた異なる条件下で進行する。
たとえば,B 細胞 ( 抗体を産生するリンパ球 ) の分化初期では,細胞はまず膜通過型ドメイン ( 細胞表面を構築する分子 ) をコードするエクソンを利用する。 その後,B 細胞は違うエクソンに切り替え,細胞から抗体分子としてのタンパク質が分泌できるようにしている。
選択的スプライシングは少数の遺伝子から多種多様のタンパク質が生産できるための機構として働く。
我々人間はわずかに約20,000の遺伝子を持っていることがわかってきたが,産生されるタンパク質の種類は少なくともその10倍にも及ぶだろう。
我々の遺伝子の92-94%が選択的スプライシングによりmRNA前駆体を生産するものと推定されている。
選択的スプライシングのパターンが組織ごとに異なって,調節されていることを示唆する証拠も得られている。しかし,転写産物のすべてが機能的であるのか,エラーが発生したものも多く含まれているのかについては不明のままである。
選択的スプライシングにより,一つの遺伝子からの異なるタンパク質が生産されるだけでなく、異なる両端の5’ 非翻訳領域 (untranslated regions,UTRs)、3’UTRs も形成される。
タンパク質の情報を持っているのはコーディング領域だけだが,その他の部分も翻訳には欠くことのできない部品である。以下に2つの例を示す。
- ショウジョウバエの中のbicoid遺伝子の3′ UTRは,胚前方にmRNAを分配する。
- アフリカツメガエルのVegT遺伝子の3′ UTRは、胚の植物極にそのmRNAを分配する。
この最も良い例は,ショウジョウバエにおける Dscam 遺伝子である。
これは単一遺伝子で,約 116 個のエクソンを含み,そのうちの 17 個が最終的に mRNA に引き継がれる。あるエクソンは常に利用され,他のものは配列から選択されて,はじめて利用される。
ショウジョウバエの細胞表面タンパク質Dscam1は,mRNA の選択的スプライシングによりおよそ19,000通りもの膨大な数のアイソフォームをつくりだすことができる。
個々のニューロンはそれぞれ異なったアイソフォームのセットを発現することにより異なる“分子標識”でラベルされており,自己と非自己の違いが生み出されている。
Dscam1のもうひとつの重要な特徴は,同じアイソフォーム同士で結合する厳密性を示すことである。
さらに,この同種の分子が結合する性質により最終的に軸索あるいは樹状突起における反発シグナルを誘起する。これにより,自己の軸索あるいは樹状突起のあいだには交差が起こらないことが保証されている。
| ヒト21番染色体上に神経接着因子をコードしている遺伝子「DSCAM」がある。この染色体を3コピーもつと,「DSCAM」の作用でダウン症の特徴的な症状を示す(正式名称,Down Syndrome Cell Adhesion Molecule の由来である)。しかし,ショウジョウバエのDscamタンパク質が働いているように,ヒト「DSCAM」はニューロンの自己認識を特殊化させていないようである。プロトカドヘリンと呼ばれている細胞表面に存在するタンパク質がその機能を果たしているようだ。プロトカドヘリンは,細胞と細胞を接着させるタンパク質であるカドヘリンのサブセットである。プロトカドヘリンは53個の遺伝子ファミリーによってコードされている。しかし,一つのニューロンではこれらのわずか6個が発現している。これら6個の遺伝子が選択されたのは偶然に依るので,各々のニューロンの自己-非自己を決定している何千もの組合せを可能としている。ショウジョウバエにおける上述した過程でのように,一つのニューロンの樹状突起が互いに遭遇した場合,同一集団のプロトカドヘリンだと反発シグナルが誘発されるので,このためシナプスが確立することはない。このように、何千もの異なるニューロンは、自己の軸索あるいは樹状突起のあいだには交差が起こらないことが保証されている。この Dscam タンパク質はニューロンが正常に発生していくのに役立っている。おそらくヒトでも,限られた数の遺伝子転写産物の選択的なスプライシングによって,信じられないほどのシナプス結合の多様性が生じているものと考えられる ( ヒトの中枢神経では ~1014 個のニューロン結合があるといわれている ) 。 |
RNA の特定の部分がエクソンとして残るのか,イントロンとして切り捨てられるのかは条件が変わることによって変化することがある。すなわち,
- 遺伝子を包含している細胞の種類による
- その細胞の分化のステージによる
- 細胞が外部からどのような信号を受け取ったかによる
選択的スプライシングに切り替えることによって,綿密な制御が成されることは明らかであろう。
トランススプライシング Trans-splicing
大部分の遺伝子は転写され,転写産物が先に述べたように処理される。RNAポリメラーゼは一つの遺伝子座位の一本鎖を下降しながらmRNA前駆体を形成する。そして,イントロンが除去され,成熟したmRNAを形成する。
しかし、例外がある。
2個の異なるmRNA前駆体間でスプライシングが起こり,本来コードされていない配列が付加される事例が多数見つかっている。
この現象は,トランススプライシングと呼ばれている。
例:異なる遺伝子座からの転写産物を連結して一つのRNA分子が合成される。
- 同一染色体だがかなり離れて位置する遺伝子
- 同一遺伝子座位の逆鎖の上遺伝子
- 相同染色体上の2つの対立遺伝子
これらのトランススプライスされた転写産物の生物学的重要性はまだ不明の点が多い。
遺伝子はなぜ分断されているのか?
恐らく進化の過程で,真核生物の遺伝子はより小さな,原始的な遺伝子(すなわち今日のエクソン ) から組み立てられてきた。
あるタンパク質は上述した 抗体 のように,いろいろ分離された領域(ドメイン)から成る情報を 1 セットもっており,これが組織化されて完成した分子となると特有の機能を発揮できる。
それぞれのドメインは分断されているエクソンにコードされている。
抗体分子のいろいろな機能を別々のエクソンによってコードしておくことによって,異なった組み合わせでこれらの単位を利用することが可能となる。
つまり,ゲノム中の 1 組のエクソンは,子供が好きなように組み立てられる 「 レゴ 」 の箱の中の様々なピース部品に相当する遺伝的なモジュールということができる。
しかし,エクソンによってはそのドメインの境界がタンパク質ドメインの境界と一致していない場合がある。したがって,分割された遺伝子の利点として,選択的スプライシングによって単一の遺伝子から多種類のタンパク質を産生できる機会が生じて来ることが挙げられる。
まとめ
遺伝子の発現は 2 つのステップで起こる。
- 転写 – DNA にコード化されている情報を RNA 分子に書き写す。
- 翻訳 – mRNA のヌクレオチドにコード化された情報をタンパク質を構成するアミノ酸の配列に置き換えていく( この詳細については 遺伝子の翻訳: RNA → タンパク質 参照 )。
真核生物では核で転写が起こり,次いで細胞質で翻訳が起こる。核をもたない細菌では,転写と翻訳が細胞質で同時に起こる。〔前核生物における遺伝子発現はこちら〕
| 最初に戻る |
| メニューのページへ戻る |
March 07, 2020