| このページの内容 |
テロメア telomere
真核生物の各 染色体 は種々のタンパク質が結合した DNA 分子 から成る。
例: 平均的なヒトの染色体は約 1 億 5 千万個の ヌクレオチド 対 ( 150 x 106 bp ) から成る DNA 分子を含む。この分子を真っ直ぐに伸ばすと,5 cm にもなる。有糸分裂 の間,染色体の長さは約 5 µm となり ( 10,000 分の 1 に短縮 ) ,DNA は極めて凝縮された形態を示す。
真核生物の DNA 分子は直線的で,両端をもつ ( これは 大腸菌 の様な細菌の染色体と著しく異なる。大腸菌では環状構造を示す ) 。
典型的な染色体の DNA 分子は,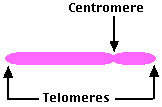
- 遺伝子 ( タンパク質や RNA をコード化している ) の直線的な配列を含む。ただしこれらは
- 何もコード化していない多量の DNA によっていくつにも分断されている。
何もコード化していない DNA 領域には,
- 動原体 centromere を構成する広範な領域と
- 染色体の末端部の広範な領域,すなわち テロメア telomere がある。
テロメアの機能
 テロメアは細胞の寿命にとって極めて重要である。また,
テロメアは細胞の寿命にとって極めて重要である。また,
ヒト ( ならびにマウス ) のテロメアは以下の塩基配列が 2000 反復もある。
5′ TTAGGG 3′.
5'...TTAGGG TTAGGG TTAGGG TTAGGG TTAGGG TTAGGG..3' 3'...AATCCC AATCCC AATCCC AATCCC AATCCC AATCCC..5'
染色体の複製
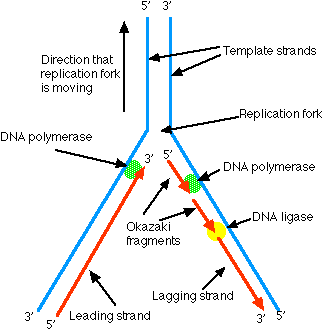
DNA ポリメラーゼがテンプレートに沿って 3′ → 5′ 方向に移動する時に,新しい DNA 鎖が合成される。染色体の 3′ → 5′ 鎖に対しては,複製起点 から中断することなく DNA ポリメラーゼが移動しながら合成していく。
しかし,5′ → 3′ 鎖をテンプレートとして合成する場合には,不連続にならざるを得ない。複製フォークが十分に開いた時に,DNA ポリメラーゼが,反対方向に移動しながら相補的な DNA 鎖の断片を合成し始める ( この断片は 岡崎フラグメント と呼ばれる ) 。その後,DNA リガーゼ が岡崎フラグメントを縫い合わせ,DNA 鎖が完成していく。
| DNA 複製についてはこちら |
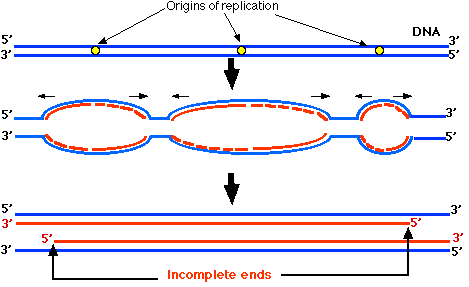
右図では,水平の黒矢印が複製フォークの移動する方向を示す。複製フォークが 3′ 方向へ移動する時はいつも新たに合成される DNA 鎖は岡崎フラグメント ( 赤波線 ) として合成が始まる。
これは染色体の末端に近づくまで継続するが,新たに合成された DNA 鎖の 5′ 末端は完全に複製されないままに終了してしまう。
したがって,それぞれの娘染色体はテロメアの部分が “短縮” した状態で複製が終わる。
ヒトのテロメアは有糸分裂毎に約 100 塩基分程の DNA が短縮すると推定されている。
このことは約 16 反復の TTAGGG が失われることを意味する。この割合でいくと,125 回の有糸分裂の後にはテロメア部分が完全になくなってしまうことになる。
これが,通常の体細胞の有糸分裂の回数に制限がある理由であろうか?
テロメアと細胞の老化
テロメアは有糸分裂毎に確実に短縮していき,これによって細胞に有限の寿命を背負わせている。これらの現象は実際に確認されている。正常な ( ガン化していない ) 細胞を培養しても,無制限には増殖しない。
| 培養された正常な細胞とガン細胞の差についてはこちら |
新生児から採取した細胞を培養した場合,およそ 100 回分裂することができる。しかし,終了前には有糸分裂速度は ( 2 週間に 1 回以下にまで ) 低下する。( 私のような ) 高齢に達した人の細胞を培養すれば,数十回の分裂を経て,細胞は分裂を停止 ( 死滅 ) するだろう。
テロメアの短縮は細胞系譜の寿命を決定する時計なのであろうか?
それを支持する観察:
細胞の中には無限の細胞分裂能をもつものがある。
- 生殖系列の細胞 ( 生殖細胞 )
- 単細胞真核生物 ( ゾウリムシなどの原生動物 )
- ある種のガン細胞
テロメラーゼ telomerase という酵素の助けを借りて,自身のテロメアの長さを維持できることが知られている。
テロメラーゼ
テロメラーゼは DNA 鎖の 5′ 末端にテロメアの反復配列を付加する酵素である。テロメラーゼ活性の高い細胞では,DNA の複製に先立ちテロメア部分を伸張せせることによって,複製時に短縮するテロメア長を補正することができる。
テロメラーゼは:
- リボヌクレオ・タンパク質 ( RNP ) ( RNA とタンパク質の複合体 ) である。
- その RNA 分子には AAUCCC (哺乳動物の場合) というテンプレートが含まれており,これにより TTAGGG が挿入される。
- すなわち,テロメラーゼは 逆転写酵素 reverse transcriptase ( RNA テンプレートから DNA を合成する酵素 ) である。
テロメラーゼは通常,以下のもので認められている:
- 生殖系列の細胞 ( 胚性幹細胞 ES 細胞を含む )
- 単細胞真核生物
- ガン細胞
しかし実験室内で,正常な体細胞にテロメラーゼを発現するよう形質転換すると,それらは通常の寿命を超えて,テロメアの短縮がないまま有糸分裂を継続する ( Bodnar ら,1998 年 1 月 16 日号 Science ) 。これらのことから,テロメラーゼとテロメアの長さが細胞の不死の鍵であることが示唆される。
テロメラーゼとガン
ほとんどのガンは体細胞から変化して,発生する。しかし,通常の体細胞とガン細胞の最も特徴的は違いは,無限に分裂する能力をもつか,もたないかである。ほとんどのガン細胞がテロメラーゼを合成する能力を取り戻しており,これによりテロメアがそれ以上短縮しないように作用していることが判明している。
恐らく,テロメラーゼの発現を防ぐ ( あるいはこの酵素の作用を防ぐ ) 薬品の投与が,ガンを撲滅するための新しい治療法となるだろう。
テロメラーゼと移植細胞
遺伝子治療の主なステップは:
- 患者から細胞を取り出す。
- その細胞に,患者が自ら合成できないタンパク質の遺伝子を導入する。
- 形質転換細胞を患者に移植する。
この手法の 1 つの問題は,すべての正常な体細胞に対しても致命的であるということである。何回かの有糸分裂の後に,細胞は死滅する。したがって,治療を受けた患者は形質転換した T 細胞を定期的に注入されなければならない。。
患者の細胞が,治療のための遺伝子ばかりでなく,活性をもつテロメラーゼ遺伝子でも形質転換された場合にはどうなるのか? これによって,無限の寿命が与えられるはずである。
しかし,ガン細胞がテロメラーゼを産生する能力を再び獲得すると,結果は逆転する。また,活性をもつテロメラーゼ遺伝子で形質転換された細胞はガン化するのだろうか?
おそらく,そうはならない。Bodnar ら ( 1998 年 1 月 16 日号 Science ) によると,そのようにした細胞はさらに 1 年間培養で発育を継続し,それらがガン化したかどうかが試験されている。
| 詳細はこちら |
結果は明るいもので,これらの細胞は分裂を継続しているが,
- それらは,正常細胞を培養した時みられるような 接触阻止現象 contact inhibition を示す。
- 免疫不全マウスに注入されても,ガン化しない ( ガン細胞を注入するとガンが生じる ) 。
- 栄養素に対する発育差を示す。すなわち,ガン細胞を維持できる単純な培地では発育しない。
- 依然として,正常な 核型 を保有している。ガン細胞では異常な核型がみられる。
テロメアとクローニング
有名なヒツジ ドリーは,成長したヒツジの体細胞から取られた細胞を,培養で増殖させた後に核移植されて作られたクローンである。細胞提供ヒツジは 6 歳であった。
| ドリーの作出についてはこちら |
ドリーのテロメアはどうなっていたのか?
ドリーの細胞のテロメア長が分析され,通常の 1 歳のヒツジにおける長さに比べると 80% しかないことが判明した。ドリーに発生した核がテロメラーゼ活性を失っていたとしても当然であろう。
胚細胞からクローニングされた他の 2 頭のヒツジもテロメアが短縮していたが,ドリー程ではなかった。おそらく,移植する前の体外培養時間の違いによるものであろう。
ドリーのテロメアが短かったといくことが,体細胞を用いた哺乳動物のクローニングにおいて新たな問題を提供している。
| 最初に戻る |
| メニューのページへ戻る |
February 08, 2020