ヒトの生殖器と生殖細胞
| このページの内容 |
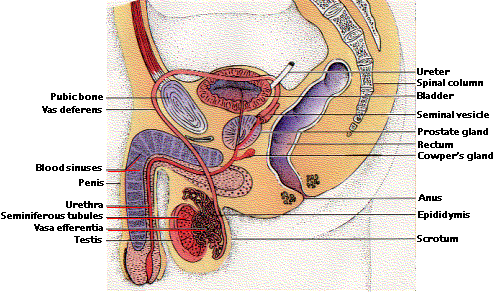
男性の生殖器
雄性生殖システムは 2 つの主な機能をもつ:
- 生殖細胞 ( 精子 ) の生産
- 受精部位への移動
精子形成 spermatogenesis は 精巣 testis 内で行われる。
| 男性生殖器の模式図はこちら |
| 家畜の生殖器のページはこちら |
{kind=link}
各精巣は 精細管 seminiferous tubule によって占められ ( ヒトの場合,引き延ばした全長は 20 メートル以上になる ),そこが精子形成の場である。
{kind=link}
精子形成
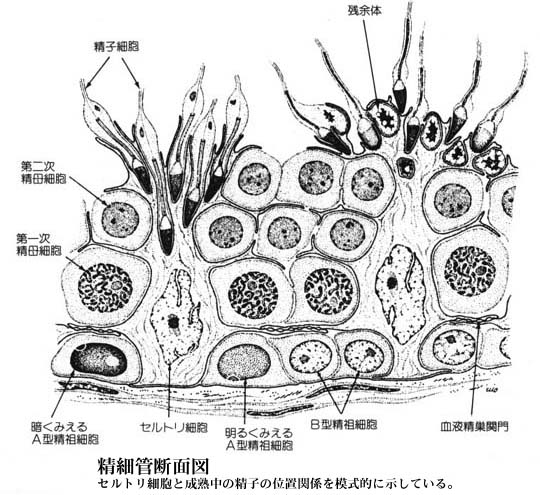
精細管の断面を見ると,精細胞と呼ばれる丸い形の細胞が幾重にも重なっている。これは精子形成に至る諸段階の細胞で,外から内に向かって進む。
精細管の管壁側には 2 倍体 の精祖細胞 spermatogonia,すなわち精子となる 幹細胞 がある。つづいて,
- 第 1 次精母細胞 primary spermatocyte
- 第 2 次精母細胞 secondary spermatocyte ( 精娘細胞ともいう )
- 精細胞 spermatid ( 精子細胞ともいう )
- 精子
の順に発育が進む。これらの細胞は,セルトリ細胞 Sertoli cell と呼ばれる背の高い細胞に接しており,栄養を供給されている。
| セルトリ細胞と成熟中の生殖細胞との位置関係についてはこちら |
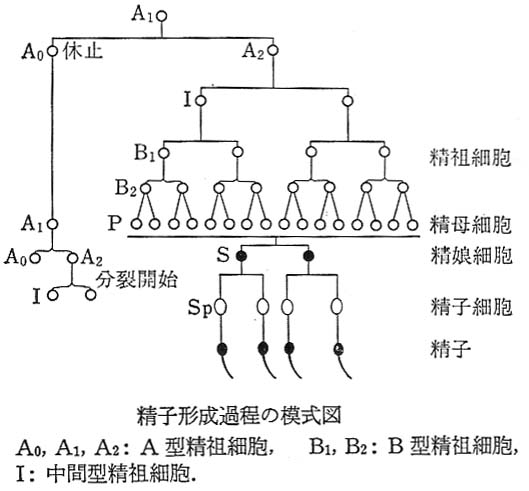
精祖細胞 は,
- 有糸分裂 mitosis によって増殖し,
- 精母細胞 spermatocyte に分化する。
各精祖細胞は 減数分裂 meiosis によって,4 個の 半数体 haploid の 精子細胞 spermatid を生産する。ヒトの場合,この過程が完了するのに 3 週間かかる。
その後,精子細胞は精子に変態する。この過程で 細胞質のほとんど が失われる。
簡単のために,図には単一の乗り換えを起こした一対の相同染色体の行動様式を示す。ヒトの場合,22 対の 常染色体 autosome がそれぞれの相同染色体間で平均 2 ヵ所の 乗り換え ( 交叉 ) を示すので,精子の遺伝子構成は極めて多様である。
| 精子形成の詳細な模式図はこちら |
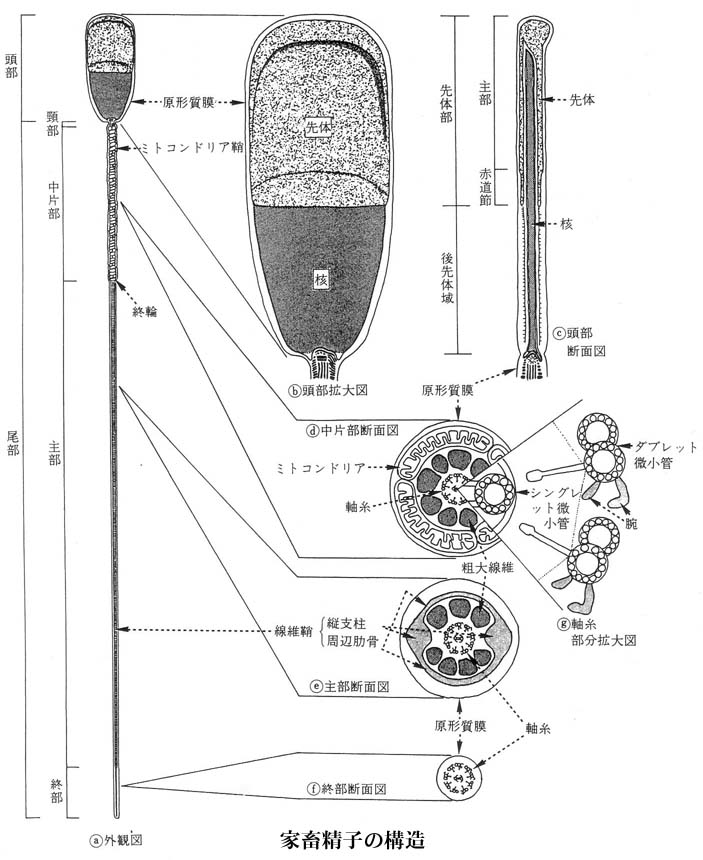
精子の構造
 精子 sperm は核に鞭毛が生えたようなもので,次の各部より構成される。
精子 sperm は核に鞭毛が生えたようなもので,次の各部より構成される。
- 頭部には
- 先端に先体 acrosome と
- 凝縮した半数体の染色体セットをもつ。
- 中片部 midpiece には ミトコンドリア mitochondria と 1 個の 中心小体 centriole がある。
- 尾部 tail
| 精子の電子顕微鏡写真はこちら |
| 家畜精子の構造についてはこちら |
{kind=link}
成人男性の場合,毎日 1 億以上の精子を生産する。これらが徐々に 精巣上体 epididymis,ついで 精管 vas deferens に移動する。ここで精子は成熟し,貯蔵される。
精子の生産の他に,精巣はホルモンを分泌する 内分泌腺 endocrine gland である。その主なホルモンは テスロステロン testosterone であり,男性の二次性徴の発達を促す。テストステロンは 精子の生産 にも必要である。
| テストステロンについて |
テストステロンは精細管の間隙を埋める 間質細胞 interstitial cells( ライディッヒ Leydig 細胞 )で生産される。
LH
間質細胞は,下垂体 pituitary gland 前葉から分泌される 黄体形成ホルモン luteinizing hormone ( LH ) の標的細胞である。
FSH
卵胞刺激ホルモン follicle-stimulating hormone は精祖細胞に直接働き,精子の生産を刺激する ( テストステロンの合成に必要な LH も働く )。 [ さらに理解するために ]
女性の生殖器
ヒトを含む哺乳動物では,生殖活動における女性の役割は男性に比べはるかに大きい。
雌性生殖システムは
- 卵子を生産し,
- 雄性生殖細胞 ( 精子 ) を受け入れ,
- 受精と 着床 の場を提供し,
- 妊娠中の胎児や分娩後の子供を育てる。
[ 模式図はこちら ]
{kind=link}
卵子形成
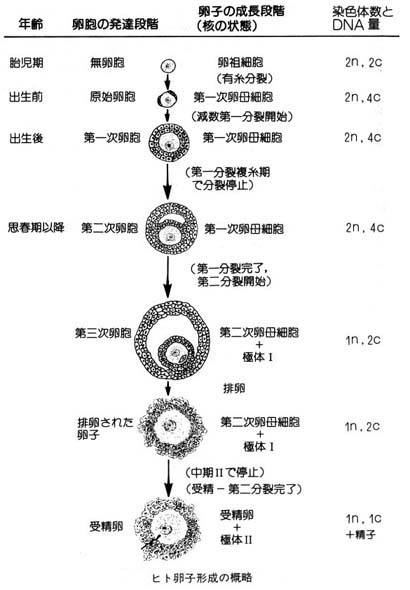
卵子形成は 卵巣 ovary で行われる。
男性と異なり, 卵子生産の最初の段階は出生前に始まる。胎児が 25 週齢になるまでに,有糸分裂 mitosis によりすべての卵祖細胞が形成される。これらの 2 倍体細胞は
- 1 次卵母細胞 primary oocytes に発育し,
- 第 1 減数分裂( meiosis I )の最初の段階を開始して,
- 停止する。
女性が性成熟に達するまで,卵子の発育はそれ以上進まない。性成熟に達して卵子の発育が再開される。
1 次卵母細胞は非常に大きく発育し,第 1 減数分裂 を完了する。そして,大型の 2 次卵母細胞 secondary oocyte と染色体の 1 セットを受け継いだ小型の第 1 極体 polar body を形成する。どちらの染色体セットが卵子内に残り,あるいは極体として放出されるのかは確率の問題である ( 第 1 極体はそれ以降の発生に関与しない ) 。
2 次卵母細胞が 第 2 減数分裂 の 中期 metaphase に達すると,再び停止する。
受精 fertilization が起こって初めて第 2 成熟分裂が完了する。精子の侵入によって,細胞周期が再開し,
- 成熟 ( または M 期 ) 促進因子 MPF ( M-phase promoting factor ) の活性が低下し,
- 細胞周期が再開する。
この第 2 成熟分裂の完了によって, 2 次卵母細胞は 第 2 極体 を放出して,受精卵,または接合子となる。
精子形成の模式図と同様に,染色体の行動様式を簡単に示してある。
 |
写真は卵子形成中の極体形成を示す。 |
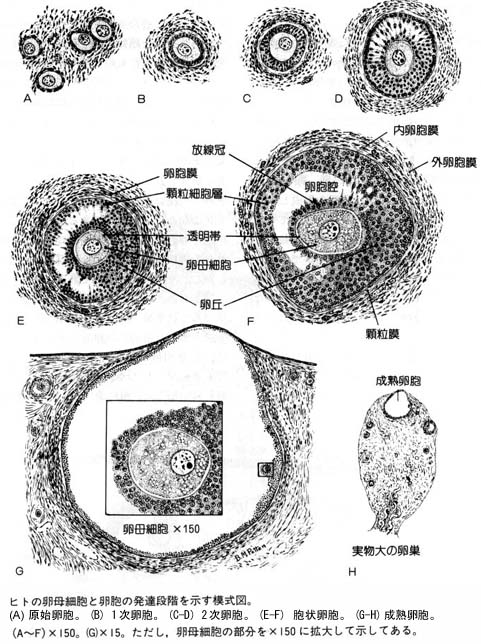
これらの現象は 卵胞 follicle 内で起こる。発育中の卵胞卵は多数の 顆粒層細胞 によって囲まれている。
成熟した卵胞は 内分泌腺 として機能する。卵胞内の細胞が エストロジェン estrogen と総称される ステロイド ホルモンを産生する。 エストロジェンは女性の二次性徴を発達させる。
- 骨盤の拡張
- 乳房の発達
- 恥毛の発生
- 女性らしい体つきとなる 脂肪組織 の発達
エストロジェンは女性の生殖期間を通して分泌される。この間,性周期 menstrual cycle に重要な役割を果たす。
| 卵子形成と卵胞発育に関する詳細な模式図はこちら |
| 卵母細胞と顆粒層細胞との関連についてはこちら |
| 性周期についてはこちら |
| ヒトの卵胞の発育についてはこちら |
{kind=link}
排卵
排卵は LH サージ に反応して起こる。排卵された 2 次卵母細胞は 卵管 fallopian tube の腹腔開口部に受け取られ,子宮に向かって移動する。
受精
受精のために,精子は体外に射出されるが,この 精管 vas deferens を通過する際に,精嚢腺 seminal vesicles, カウパー腺 Cowper’s glands,ならびに 前立腺 prostate gland の分泌液が加わる。 [ 生殖器参照 ]
これらの分泌液は精子にとってエネルギー源 ( フラクトース fructose ) となり,化学的に最適な環境となる。
| 家畜の副生殖腺についてはこちら |
精液 semen は精子と副生殖腺の分泌液から成る。尿道 urethra を通り,射出される。
射出された精子は膣,子宮を通過し,受精部位である卵管に達する。卵管にある卵子 ( 2 次卵母細胞 ) と会合すると受精が起こる。
精子自体は運動性をもつが,受精部位までの上走には子宮や卵管の筋運動が関与している。射出時には数億の精子が含まれるが,受精部位に到達できるのは数百といわれている。そしてこれらのうち,1 個の精子だけが卵と受精する。
受精時には,
- 卵を被う 透明帯 zona pellucida に精子頭部が結合する。
- 精子頭部の先端にある 先体 acrosome から
- エキソサイトーシス exocytosis 現象によって,透明帯を通過するために必要な 酵素 が放出され,
- 精子頭部は卵の細胞膜と結合することができる。
- 両者の細胞膜同士が融合し始めると,精子全体が卵細胞質に取り込まれる。
- 精子のミトコンドリアも卵の中に取り込まれるが,発生の過程でこれらは消失し,卵の遺伝子には関与しない。
したがって,ミトコンドリア DNA は常に母性遺伝する。
- 精子侵入と同時に, 卵子から分泌される酵素が透明帯に作用して,透明帯に達した他の精子が通り抜けられないようにする ( 多精拒否 ) 。
- 侵入精子の頭部はまもなく膨化し,雄性前核 male pronucleus に変わる。
- これと同時に,卵子 ( 2 次卵母細胞 ) は第 2 減数分裂を再開し,第 2 極体 second polar body を放出して,減数分裂が完了し,
- 雌性前核 female pronucleus に変わる。
- 雄性前核 と 雌性前核 はお互いに近づき,融合して,完全な 2 倍体の染色体構成が完成する。
- 核膜が崩壊し,精子の中心小体が複製された後,紡錘体が形成される。
- この後,受精卵 または 接合子 zygote は第 1 有糸分裂 ( すなわち 卵割 ) を開始する。
妊娠
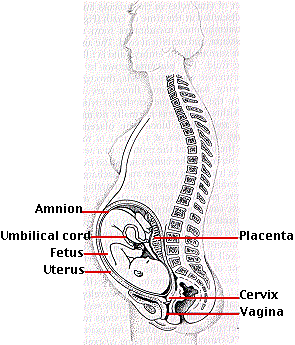
接合子が 卵管 fallopian tube 内にある時期に 卵割 が開始する。胚は分裂しながら卵管を下降し,3 日程度で子宮に達する。この間卵割が進行し,胚盤胞 blastocyst と呼ばれる胞胚腔をもった胚に発育する。受精約 1 週間後に,胚盤胞は肥厚した子宮壁に埋まり込んでいく。この過程を 着床 implantation と呼び,妊娠が成立する。
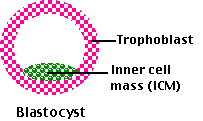 胚盤胞は2つの細胞系譜をもつ:
胚盤胞は2つの細胞系譜をもつ:
- 内細胞塊 inner cell mass ( ICM ) – 将来,胎児に発生する。
- 栄養膜 trophoblast – 胎盤等を形成する。 4 種類の 胚体外膜 extraembryonic membrane に発生し,出産までの胎児の発育中に極めて重要な役割を果たす。
胚体外膜からは 羊膜 amnion, 胎盤 placenta ならびに 臍帯 umbilical cord などが形成される。 胎盤は子宮壁に付着した状態で発育し,胎児血は絨毛表面を介して母体血と接しているが,両者は混じり合うことはない。胎盤は胎児と母体の間にあって物質輸送の中心的な役割を果たしている。
- 糖質,アミノ酸,塩類,水分などの栄養分を受けとる。
- 酸素を受けとり,二酸化炭素を放出している。
- 尿や他の老廃物を排出する。
- 出生直後の新生児免疫能に重要な役割を果たす 抗体 antibody ( 主として IgG ) を受けとる。
胎盤は物質の輸送だけではなく,妊娠の維持,胎児の発育に必要な諸種のホルモン,例えば絨毛性ゴナドトロピン,胎盤性ラクトゲン ( 略称 PL ) ,種々のステロイドホルモンを分泌している。また,多種の代謝機構をもっており,多くの酵素が存在している。
| 胎盤の内分泌機能についてはこちら |
胎盤の代謝能は胎児のそれに匹敵するほど高い。
臍帯 umbilical cord が胎児と胎盤を連絡する。胎児血は胎児内腸骨動脈に連続する 2 本の臍動脈によって胎盤に入り, 1 本の臍静脈を通って胎児へと運ばれる。
多量の酸素と養分を含んだ動脈血は, 1 本の臍静脈に集められ,臍帯を経て体内に入り肝臓や下大静脈に運ばれる。
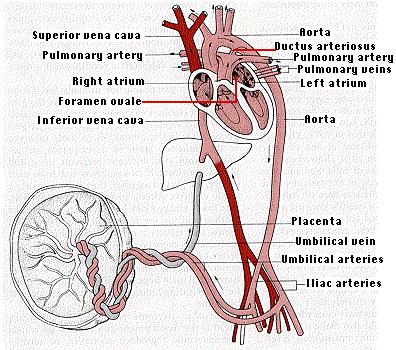 肺呼吸の行われていない胎生期の血液循環は,出生後の循環と比べきわめて特異的である。胎生期では,右心室の血液の一部が心房中隔の 卵円孔 foramen ovale を通って左心房に入り,左心室を経て大動脈に入る。
肺呼吸の行われていない胎生期の血液循環は,出生後の循環と比べきわめて特異的である。胎生期では,右心室の血液の一部が心房中隔の 卵円孔 foramen ovale を通って左心房に入り,左心室を経て大動脈に入る。
胎盤には母体血,胎児血がそれぞれ流入,流出するが,母体血と胎児血とが直接混じり合うことはない。しかし,胎児の血球が母胎血流に入り込むことがある。このため,羊水穿刺や絨毛生検などの方法に変わって,母体血を採取して遺伝病の 出生前診断 prenatal diagnosis をする方法がある。
極めてまれなことではあるが,母体血が胎児へ移行することがある。白血病やリンパ腫の女性が妊娠し,胎児へ悪性腫瘍が転移した例がある。
妊娠の最初の 2 か月に,胎児 ( 胎芽 ) の基本的な構造は形成されている。あらゆる種類の細胞への分化が起こっている。この期間に,発育中の胎児は,発育に影響するすべてのものに非常に敏感である。
母親へのウィルス伝染,例えば風疹ウィルス,あるいはある化学薬品への接触によって胎児に奇形を引き起こすことがある。そのような因子は 催奇物質 teratogen と呼ばれる。1954 年から 1962 年にヨーロッパにおいて,多くの妊娠女性に処方された精神安定剤 ( サリドマイド ) が催奇物質であることが判明し,数千人規模の不幸な事例があった。
妊娠3ヵ月までに主要臓器の発生は終わるので,これ以降は基本的に成長の期間と小さな形態変化の時期である。この期間の胎芽が最も催奇物質に対して敏感である。
妊娠は ホルモン の複雑な支配を受ける。これについては,別に述べる。 [ 妊娠のホルモン支配 ]
胎盤は 移植臓器 と似ている
「 なぜ胎盤が母体の免疫系に拒絶されることなく,9 ヵ月間も機能するのか? 」 これが免疫学における未解明の問題の 1 つである。胎盤のすべての細胞は父親のゲノム ( 父親の半数体セット )をもっている。これには第 6 染色体にある主要組織適合抗原に対する遺伝子も含まれている。
| 1 つの例外 : 父親の X 染色体上の遺伝子は 1 つも発現しない。 X 染色体の不活化が胎子の細胞で無作為に起こるが,栄養膜の細胞では無作為ではない。栄養膜 とその後継細胞では常に父親の X 染色体が 不活化 される。 [ X 染色体の不活化についてはこちら ] しかしこれで問題が解決したわけではない。というのは主要組織特異的抗原のすべてが第 6 染色体に位置し,これは不活化されないからである。 |
| 主要組織特異的抗原複合体 ( MHC ) についてはこちら |
このように,胎盤は母親にとって免疫学的な異物なのである。
しかし,胎盤は機能する。
半世紀にも及ぶ研究にもかかわらず,この免疫学的に特異的な現象の機構は未解明のままである。しかし, 1 つは確かである。
母親は,父親の抗原に本質的に寛容ではない。 その理由:
- 母親は,速やかに父親からの皮膚移植を拒絶するだろう。
- 母親は,胎児に発現した父親の組織適合性抗原に対する抗体を産生する。実際,同じ父親によって数人の子供を生んだ女性からは組織適合性試験に使用されるヒト白血球抗原の抗血清がよく得られている。
それではこの現象はどう説明できるのか? いくつかの可能性として:
- 胎盤はクラスII の組織適合抗原を発現しない。
- クラスIの組織適合抗原が非常に弱い。
- 胎盤の細胞は プロジェステロン を分泌する。このホルモンは拒絶反応を抑制する。
- ラットでは,胚 ( ならびに子宮内膜 ) が 副腎皮質刺激ホルモン分泌促進ホルモン ( CRH ) を分泌する。このホルモンは,胎盤の細胞上で Fas リガンド ( Fas ligand, FasL ) の発現を誘導する。活性 T 細胞は Fas を発現する。したがって,脅威となる T 細胞が,標的細胞上の FasL と遭遇してアポトーシス ( 細胞死 ) を起こしてしまう。
アポトーシスにおける Fas と FasL の詳細についてはこちら ( ただし,細胞障害性 T 細胞自身は FasL を有し,Fas をもつ標的細胞を死に至らしめる例が説明されている。) - マウスでは,胎盤の細胞がアミノ酸 トリプトファン を変性させる。トリプトファンは T 細胞の機能に必要である。マウスをトリプトファン変性酵素の抑制剤で処理すると,その胎子は母体のリンパ球の作用によって流産してしまう ( Munn ら, 1998, Science, 281: 1191. )
{kind=link}
分娩
 分娩の開始が何によってもたらされるのか正確なところはまだ十分に理解されていない。おそらく,多種多様のホルモンが関わり合って起こる。
分娩の開始が何によってもたらされるのか正確なところはまだ十分に理解されていない。おそらく,多種多様のホルモンが関わり合って起こる。
| 出産と泌乳に関わるホルモンの詳細はこちら |
分娩の最初の兆候は子宮頸管の開大である。強力な子宮筋の収縮に伴い,羊膜が破れて羊水が産道に流れ出し,胎児が娩出される。
出生を機にまず胎盤との連絡が絶たれ,次いで肺呼吸が開始されるとともに卵円孔は閉じ,臍動脈,臍静脈も閉塞し,新生児の血液循環路は成人にみられるものと同じパターンの肺呼吸型循環系に一気に切り替えられてゆく。
ある新生児では,この切り替えがうまくいかず,肺動脈の血流が不十分なことがある。この原因として,一酸化窒素 nitric oxide ( NO ) が十分に合成されないことが考えられている。
胎子娩出の後に,胎盤と臍帯の残部( 後産 )が排出される。
出産時ならびにその後数日間の母乳は 初乳 colostrum と呼ばれ,新生児に受動の免疫を供給する抗体を含む,カロリーおよび蛋白質に富んだものである。
分娩後 3,4 日して,乳汁分泌が盛んになる。
- 乳汁の合成は下垂体ホルモン プロラクチン prolactin ( PRL ) によって刺激される。
- 新生児の哺育が始まると,オキシトシン oxytocin 濃度が上昇し,これによってプロラクチン分泌が促進される。
- 乳汁には乳生産を抑制するペプチドが含まれており,乳腺に乳汁が残っていると乳生産が始まらない。この 自己分泌 autocrine 作用により,適切な乳汁生産が行われる。
胎児発生に障害を及ぼす物質
催奇物質 teratogen とは,胎児( 胎芽 )に異常を引き起こす物理化学的変化を生じさせるものをいう。
これらの物質による胎児奇形は非遺伝性の変化である。1960 年代には 5 種類程度の催奇物質が知られていたが,現在ではその種類ははるかに増大している。代表的なものを表にまとめた。
| 化学合成物とヒトの生殖についてはこちら |

| 最初に戻る |
| メニューのページへ戻る |
February 06, 2020