受精,卵割ならびに着床
| このページの内容 |
受精
受精 fertilization は 精子 と 卵子 の遭遇に始まり,両配偶子の核の合体で完了する。受精の完了は新しい胚発生の出発点でもある。
1. 精子と卵子の遭遇
受精が成立するためには,
- 受精能を有する精子と卵子が同じ時期に,同じ場所に存在すること
- 受精の場の環境が受精成立に適していること
の条件が必要である。
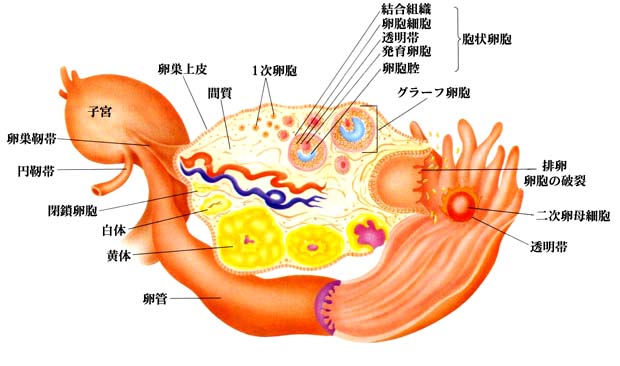
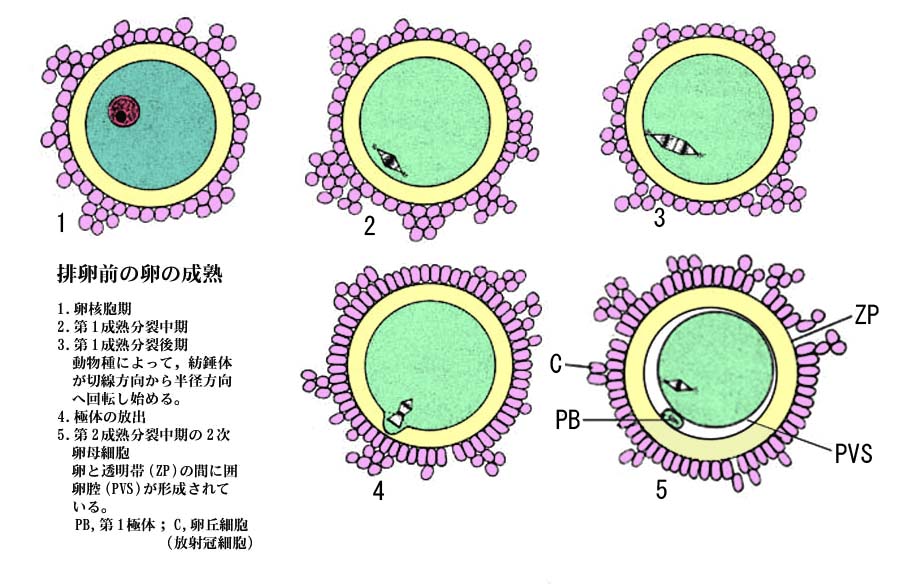
| 卵の成熟過程を示す模式図は こちら |
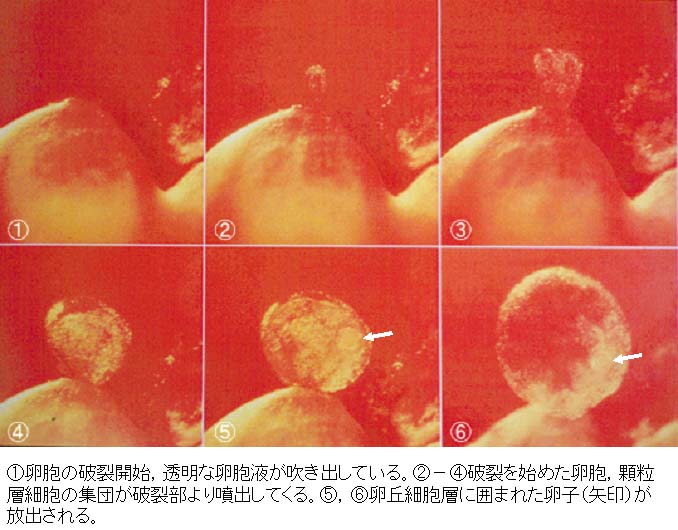
| ウサギの排卵の過程は こちら |
a) 排卵と交配との関係
周期的に排卵が起こる “自然排卵動物” では,雄を許容する発情は常に排卵に先立って始まる。
つまり自然条件下では,精子は卵子より前に雌性生殖道内に送り込まれることになる。
受精の前に,以下の準備が必要となる:
- 受精部位への精子の移送
- 雌性生殖器内で精子に生ずる機能的変化,すなわち “受精能獲得” の完了
b) 受精部位と排卵された卵子の移動
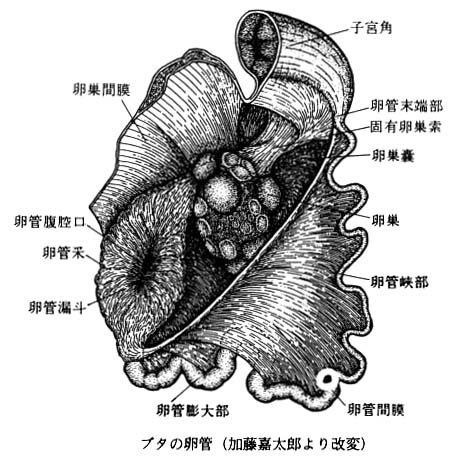
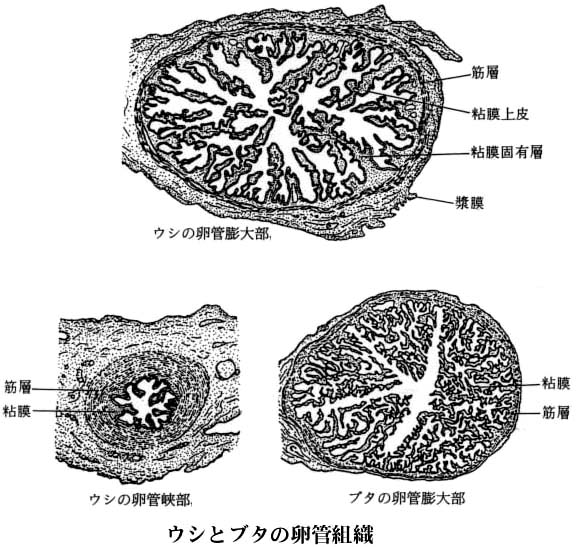
ほとんどの哺乳動物において,受精は卵管の上部,すなわち 卵管膨大部 で起こる。
卵巣と卵管の間に直接のつながりはないが,動物によって解剖学的な特徴が異なる:
- げっ歯類 や イヌ – 卵管間膜が延長して,ほぼ完全に卵巣を包んで卵巣嚢 ( らんそうのう ) を形成する。
- 有蹄類 や ウサギ – 卵管腹腔口が漏斗状に広がって,卵巣表面を覆う。
そのため,
- 粘稠な基質に包まれた排卵直後の卵子は卵管上皮の繊毛運動に送られて卵管内へ移行する。[ビデオ付き論文はこちらFigure 2を参照]
- 卵管内に取り込まれた卵子は上皮の繊毛運動と卵管壁の筋運動によって膨大部に運ばれる。
- この時期,卵管峡部との境界は非常に狭くなっているために,卵子は膨大部に留まり精子の到着を待つ。
| 齧歯類,ウサギ,ヒトの卵管の模式図は こちら |
| ブタの卵管の模式図は こちら |
| ウシとブタの卵管組織図は こちら |
c) 受精部位への精子の移動
交配によって,
- 膣内 – ウシ,ヒツジ,ヤギ,ウサギ
- 子宮頚管内 – ウマ,ブタ,げっ歯類
に射出された精子は,雌性生殖道の運動と精子自体の運動との両者によって卵管膨大部に到達する。
①精子の急速移送
精子は雌性生殖器道の筋運動によって急速に,かつ受動的に受精部位に運ばれる場合がある。
雌ウシで,交配後わずか 2.5 分の卵管上部に精子を見いだしたとの研究に基づいている。
その運搬に関わる因子として エストロジェン と オキシトシン,さらには プロスタグランジン の促進的役割が考えられる。
②卵子と精子の同時到達
上述のように,交配後直ちに卵管に達する精子群が観察されるが,これらの精子は受精部位を素通りして腹腔に達してしまう。
それに次いで卵管峡部で一時蓄えられ,受精部位に卵子が下降してくる時期に精子が卵子の近傍に達し,これらが受精に関与する。
|
| 精子と卵子の移送のまとめ |
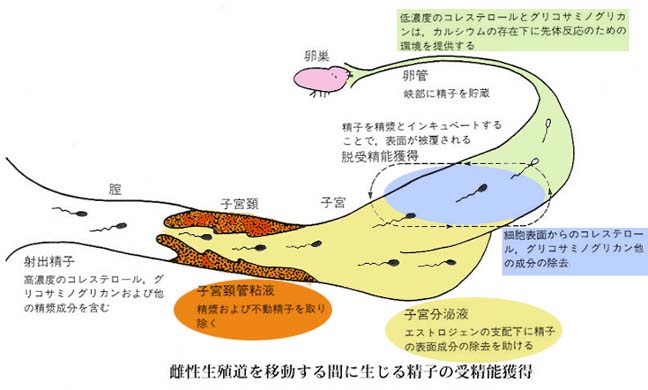
2. 精子の受精能獲得
 精子は雌性生殖道内で一定時間滞在して,生理的ならびに形態学的変化を遂げた後に,初めて受精可能となる。
精子は雌性生殖道内で一定時間滞在して,生理的ならびに形態学的変化を遂げた後に,初めて受精可能となる。
この変化を精子の 受精能獲得 capacitation と呼び,哺乳動物に特有の現象である。
| 精子の受精能獲得の最初の証拠は,1951 年に Austin と Chang によってそれぞれラットおよびウサギを用いて独自に発見された。 |
- 受精能獲得に伴って精子に生ずる変化
- 前述した発見に続き,一度受精能を獲得した精子を精漿に浮遊させると受精能を失うこと,
- また,このような精子でも雌性生殖道内に戻されれば再び受精能力を現わすこと
が観察された。これらのことから,精漿によってもたらされた変化は受精能獲得の逆の現象,すなわち 脱受精能獲得 decapacitation と考えられる。
したがって,精漿には受精能獲得の抑制因子,すなわち 脱受精能獲得因子 decapacitation factor ( DF ) の存在が予測され,受精能獲得の過程ではこの精子の表面に付着している “DF” が除去されるものと予想される。
受精能獲得( または 先体反応 )に伴い精子運動性にも著しい変化が生じ,振幅の大きな激しい尾部の鞭打ち運動を特徴とする非前進性の運動,すなわち 超活性化運動 hyperactivated movement が現われる。
また,先体 内に含まれる酵素も受精能獲得の過程で活性化される。
- 受精能獲得の誘起要因
- 体内環境( 卵管と子宮 )における誘起 – 生体内受精
- 体外における種々の処理による誘起 – 体外受精
| 精子の卵子への接近のまとめ |
3. 受精の過程
 受精能力を備えた精子と卵子がタイミング良く出会うと,いよいよ受精が始まる。
受精能力を備えた精子と卵子がタイミング良く出会うと,いよいよ受精が始まる。
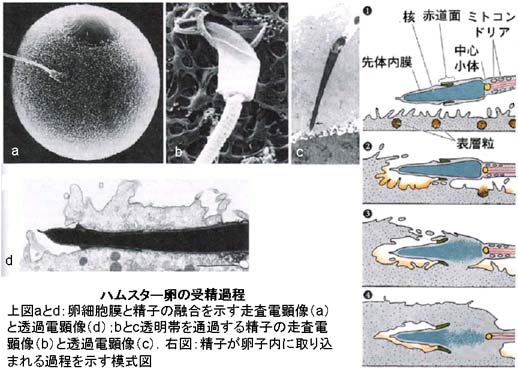
a) 卵子内への精子の侵入
- 受精能獲得精子が卵子の近傍に達すると,卵子に付随する物質に反応して精子の “先体” に変化が生ずる。
この変化は 先体反応 acrosome reaction と呼ばれ,精子頭部の細胞膜と先体外膜が部分的に融合して胞状化し,先体 の内容物( ヒアルロニダーゼを含む種々の加水分解酵素など )が放出される現象である。精子は先体内酵素の働きで,卵子をとりまく卵丘基質を溶解して,卵子に接近する。 - 精子は透明帯表面に達すると,その精子受容体( 1 種の糖蛋白質と考えられている )と結合する。
精子と透明帯,精子と卵子の結合を示す模式図はこちら - 次いで,先体内膜に含まれるタンパク質分解酵素( アクロシン )の作用と,激しい精子尾部の推進運動(超活性化)によって透明帯を通過する。
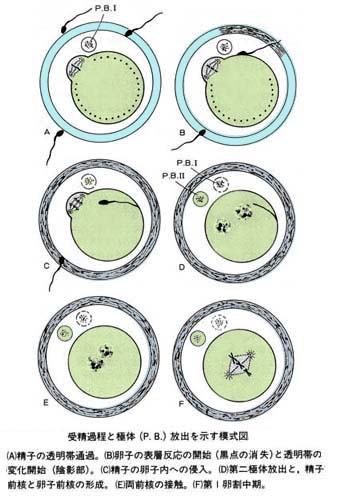
- 囲卵腔に達した精子は,頭部細胞膜と卵子細胞膜との融合によって,まず頭部が,そして精子全体が取り込まる。
- 卵子内に精子が侵入すると,卵子内では中断されていた第 2 成熟分裂が再開し,後期から終期を経て第 2 極体が放出される。
- また,表層顆粒の膜と卵子の細胞膜との融合が起こり,表層顆粒の内容物が囲卵腔に放出される。
表層顆粒から放出された物質の作用で透明帯の性質が変化し,後続の精子の結合および侵入が阻止される。これを 透明帯反応 zona reaction という 。
同時に,受精に伴う卵細胞質表面の変化 ( 卵黄遮断 vitelline block という )にも関与していると思われる。
| 先体反応のまとめ |
| 精子の侵入後に起こる変化のまとめ |
b) 前核の形成と合体
- 受精して,取り込まれた精子頭部内容物は卵子細胞質内で膨潤し,雄性前核 male pronucleus を形成する。
- 一方,成熟分裂の完了によって卵細胞内に残った半数体の染色体から 雌性前核 female pronucleus が形成される。
- その後,両前核は融合し,精子および卵子由来の染色体群の出現を経て第 1 卵割の前期へ移行する( 受精の完了 )。
4. 多精拒否
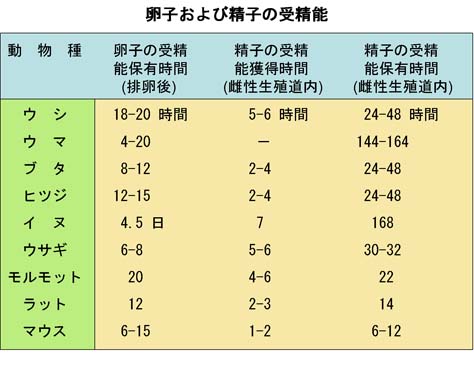
卵子は,前述したように透明帯反応と卵黄遮断によって 2 重に 多精受精 から保護されている。
動物種によって,
- 透明帯反応を主体とする型 – ハムスター,ウシ,ブタ,ヒツジ
- 卵黄遮断の強力な型 – ウサギ
- 両者の中間型 – マウス,ラット
5. 卵子の受精能力
排卵後の卵子の受精能保有時間は動物により, 6 – 8 時間( ウサギ )から約 108 時間( イヌ )までの変異があるが,ほとんどの動物種ではおよそ 24 時間以内と推定されている(右表)。
6. 異常な受精の様式
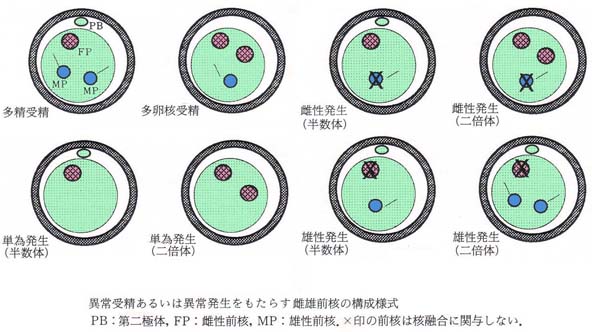
- 多精受精 polyspermy
2 個以上の雄性前核と 1 個の雌性前核が融合する現象 - 多卵核受精 polygyny
2 個以上の卵子由来の前核と 1 個の雄性前核が融合する現象 - 単為発生 parthenogenesis
精子の侵入を受けないで,雌性前核のみで発生が進む現象
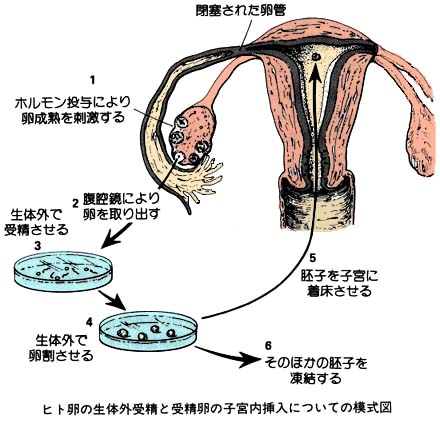
| ヒトの体外受精の模式図は こちら |
初期胚発生と着床
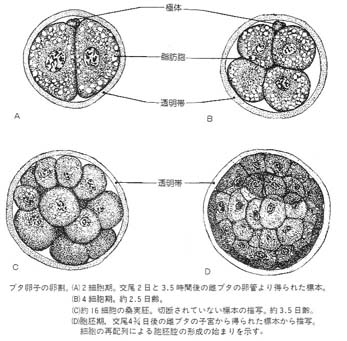
卵割 cleavage
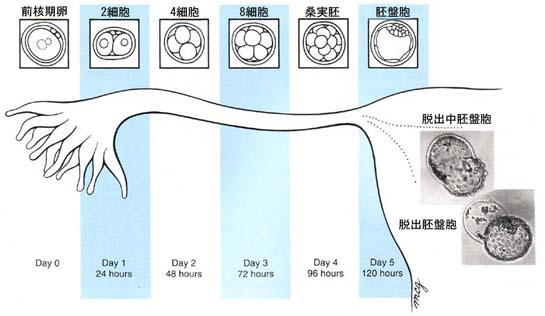 接合体 zygote が形成された後,胚は 有糸分裂 mitosis を起こす。接合体 ( 1 細胞期の胚) は通常の体細胞に比べると極めて大型で,細胞質に対する核の比率が低い。
接合体 zygote が形成された後,胚は 有糸分裂 mitosis を起こす。接合体 ( 1 細胞期の胚) は通常の体細胞に比べると極めて大型で,細胞質に対する核の比率が低い。
この有糸分裂の間は,細胞の大きさに増加は見られず,最終的に体細胞と同じような核と細胞質の比率となる。この過程を 卵割 cleavage と呼んでいる。
哺乳類の胚の細胞はほとんど卵黄を含んでいないため,妊娠初期には代謝を維持するための栄養素の大部分を母体に依存している。これらは,卵管および子宮の分泌物によって供給される( 組織栄養 )。
卵割の初期には,代謝速度はほとんど増加しないが,桑実胚から胚盤胞期に急激な上昇が起こる。
1. 卵割面
接合体の第 1 卵割面は,卵の動物極( 極体 が放出される部位 )から植物極( 卵黄に富む部位-哺乳動物の場合は明確ではありません )へ垂直に形成される。
卵割の結果, 2 つの同じ大きさの娘細胞を生ずる。これを 割球 blastomere とよぶ。
第 2 卵割では, 2 つの割球のうち 1 つは第 1 卵割と同様に割球の経線に沿って起こり,他の 1 つは水平に分裂する。
| 卵割面の方向性の違いについてはこちら |
すべての割球が同時に分裂するわけではなく,奇数の割球が観察されることがしばしばある。
卵割は,染色体の完全な組み合わせを受け継ぐ娘細胞( 割球 )を形成する有糸分裂である。哺乳類および他の脊椎動物では,初期胚の割球の運命が下等な動物種ほど強固に決定されていない。
このため, 2 細胞期胚の割球がそれぞれ 2 頭の健康な産子に発生する可能性を有している。一方,決定的な卵割予定を有する下等動物では,このような操作を施すと,異常な胚または部分的に形成された胚を生ずる。
極めて初期の割球は 全能性 totipotency,すなわち, 1 つの完全な胚になる能力を有する。しかし,割球は桑実胚内における位置によって分化し始める。
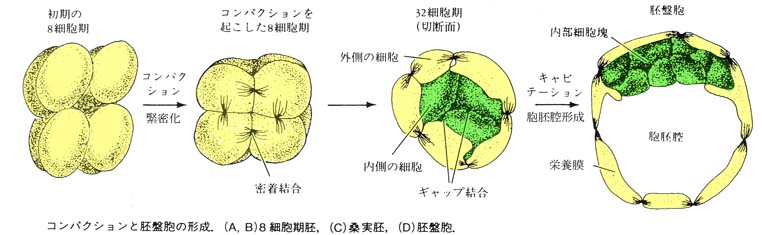
2. 胚の緊密化( コンパクション ) compaction
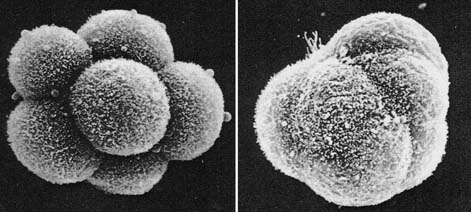 マウス初期胚では, 8 細胞期( 第 3 卵割直後 )に形態学的に特徴的な変化が起こる( マウスでは 8 細胞期だが,家畜胚ではそれ以上の細胞期でみられる )。
マウス初期胚では, 8 細胞期( 第 3 卵割直後 )に形態学的に特徴的な変化が起こる( マウスでは 8 細胞期だが,家畜胚ではそれ以上の細胞期でみられる )。
初め 8 細胞期胚の割球同士の接着は緩い配列をとっているが,しばらくして割球はお互いに扁平になりながら密着して,胚全体がほぼ球形の胚となる。この現象を 緊密化 ( コンパクション compaction ) と呼ぶ。
緊密化の特徴である強い細胞間結合,すなわち密着結合 tight junction が球形の胚を構成する外側の細胞間で形成される。
胚のすべての割球は ギャップ結合 gap junction で連結され,隣接した細胞間では小分子やイオンが通過できる。 緊密化を起こした胚の細胞は,分裂して 16 細胞期の 桑実胚 morula をつくる(マウスの場合)。
桑実胚期には,割球による液体の分泌によって,胚内部に胞胚腔 blastocoel ができる。胞胚腔が形成された胚は胚盤胞 blastocyst と呼ばれる。
胚盤胞の外側を構成する細胞は,栄養膜 trophoblast と呼ばれ,この細胞からは胚( 胚子 )は形成されず,胎子胎盤の一部である 絨毛膜 chorion 《胎子が母体から酸素や栄養分を得るための組織で,母体の子宮が胎子を保持するためのホルモンを分泌する組織》が生じる。
また,母親が移植された器官のように胎子を拒否しないよう免疫反応の調節因子を作る組織でもある。これら栄養膜は胚が子宮に 着床 implantation する時に必要である。
栄養膜に囲まれた内側の細胞集団を 内細胞塊 inner cell mass ( ICM ) という。
胚( 胚子 )はこの内側の内細胞塊から発生する。
このように,栄養膜と内細胞塊の割球の分岐は,哺乳類の発生における最初の分化である。
【図の説明】初めのうちは,桑実胚に腔所は見られない。しかし,胞胚腔形成 cavitation と呼ばれる過程で,栄養膜の細胞は桑実胚内部に液体を分泌し,それが貯留し 胞胚腔 blastocoel を形成する。内細胞塊は,球形の栄養膜の内側に位置する。この構造を 胚盤胞 blastocyst と呼ぶ。
3. 割球の極性化
哺乳類の発生における最初の分化,すなわち内部細胞塊と栄養膜への分離が生じるための状況は,胚の緊密化によって起こる。
最近,ウシ胚では割球表面の微絨毛の分布を指標とすると,9 – 16 細胞期に 極性化 polarization を示す割球が出現し始め,16 細胞期以降はほぼ半数 ( 42% ) の割球が極性を有することが明らかにされている。この割球の極性化と核の全能性との関連が検討されはじめている。
受精卵の移動と着床
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. 受精卵の卵管内移動
排卵卵子が卵管膨大部に達するのは比較的短時間であるが,受精した後卵管から子宮に進入するまでに要する時間は,小数例を除いておよそ 3日前後である。
この間,卵管内卵子は 膨大部 – 峡部接合部 ampullary-isthmic junction ( AIJ ) 付近に一定時間滞留後,卵管峡部に侵入する。
子宮-卵管接合部 uterotubal junction ( UTJ ) には卵子移動の停滞を招くような特別な構造はみられず,AIJ または峡部全体が卵子移動を規制しているようである。
卵子移動の要因としては,
- 卵管上皮細胞の繊毛運動と卵管内粘液流動,
- 卵管輪状筋の蠕動性収縮運動
などがあげられる。 受精卵は卵管内移動中も発生を続け,子宮へ移行する。
子宮内進入時期の卵割期は下表の通り。
| 子宮内進入時期の卵割期 | |
| ブタ | 4 細胞期 |
| マウス,ハムスター,モルモット | 8 細胞期 |
| ラット,ヒツジ,ウシ | 8 – 16 細胞期 |
| ウマ,アカゲザル | 16 細胞期 |
| イヌ,フェレット | 16 – 32 細胞期 |
| ウサギ | 桑実期 |
| ネコ | 後期桑実期 |
2. 胚の子宮内分布
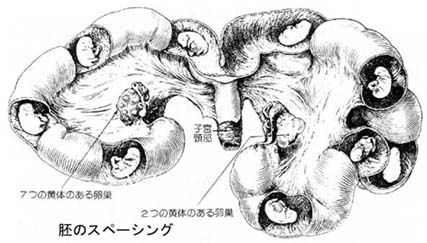 多胎動物において,卵管より子宮角内に進入した胚は,最初子宮角の先端付近に集団で位置しているが,時間の経過とともに次第に子宮頚方向に移動し,相互に一定の間隔を保ちながら子宮角内に分布していく。
多胎動物において,卵管より子宮角内に進入した胚は,最初子宮角の先端付近に集団で位置しているが,時間の経過とともに次第に子宮頚方向に移動し,相互に一定の間隔を保ちながら子宮角内に分布していく。
多胎動物におけるこのような現象を スペーシング ( spacing,間隔どり )という。
また,胚は通常排卵側卵管から子宮内に進入するが,子宮体を経て反対側子宮角内に着床するものも,ブタなどで多く観察されている。
この現象を胚の 子宮内移行 transuterine migration とよぶ。胚の子宮内移行の発生率は,ウシ,ヒツジで低く,ウマで高い。
3. 着床
着床の様式や過程は動物種によりかなり変異があるが,子宮内胚は:
- 胚盤胞 blastocyst
- 拡大成長した 拡張胚盤胞 expanded blastocyst
- 透明帯を破って脱出中の 脱出中胚盤胞 hatching blastocyst
- 透明帯から脱出した 脱出胚盤胞 hatched blastocyst
を経て,発育が継続する。
 このように胚は子宮内進入後ただちに着床することなく,動物種により定まった期間子宮腔内を浮遊している。
このように胚は子宮内進入後ただちに着床することなく,動物種により定まった期間子宮腔内を浮遊している。
その後,子宮壁の一定部位に定着して胚の発育の準備を始めることを 着床 implantation という。
より厳密には,胚盤胞の外壁を成す栄養膜が子宮上皮細胞と接着した時点から,内膜侵襲の時期までを含めて定義される。
a. 胚盤胞の定位
拡張した胚盤胞は子宮壁に接してやがて定着するが,子宮に対する胚盤胞の着床部位と方向が決まることを 定位 orientation という。
胚盤胞の定位は子宮壁に対する 胚盤 germinal disk の位置によって,
- 間膜側mesometrial,
- 反間膜側 antimesometrial,
- 側位 lateral
に分けられる。
この定位によって,着床後の胎膜や胎盤の形成が異なってくる。胚の定位を決定する本質的な因子はまだ明らかではない。
b. 着床の様式
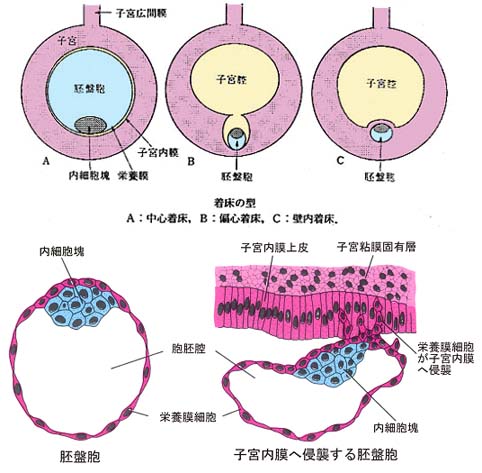
胚盤胞の表面にある栄養膜と,母体の子宮内膜の接着様式には,大別して次の 3 通りがある:
- 中心着床 central implantation
胚盤胞は子宮内に拡張して,栄養膜が子宮内膜全面的に接する。なお,栄養膜が子宮内膜の内部に侵入しないものを 表面着床 superficial implantation ともいう。 一般に,家畜胚の着床はこの様式が多い。
ウサギの胚盤胞は,はじめ中心着床( 表面着床性 )であるが,しだいに偏心着床のげっ歯類のような胎盤形成をする。 - 偏心着床 eccentric implantation
胚盤胞が子宮内の片側( 反間膜側 )に寄り,子宮内膜の襞の窪みにおさまり発育し,子宮腔の中心より離れて偏在して着床する型である。多くのげっ歯類がこの様式をとる。 - 壁内着床 interstitial implantation
子宮内壁の一側に胚盤胞が子宮内膜上皮を通って侵入し,そこに埋まって着床する型である。ヒトを含む高等霊長類,モルモット,ハリネズミなどの食虫類がこの様式をとる。
| 最初に戻る |
| メニューのページへ戻る |
February 06, 2020