化学合成物とヒトの生殖
化学合成物はどのようにヒトの生殖能力を変えてしまうのか?
| このページの内容 |
生殖システムが脅かされている
>環境ホルモン ここ数年,化学合成物,医薬品,溶剤ならびに環境汚染物質がステロイドホルモン( エストロジェン や アンドロジェン )の類似効果あるいは抑制効果を示すことが指摘されている。ホルモンは,生殖システム,細胞の恒常性の維持や体の発育に重要な働きを持つ。
ある研究によると,過去 50 年の間に男性の生殖機能が変化しており,とくに 精子 の数と質の低下,生殖器の奇形率や精巣ガン発生率の増加が示唆されている。
 以下の物質は,ヒトの受精能力に影響し,精巣ガンおよび乳ガンの発生率を増加させると疑われている物質である。
以下の物質は,ヒトの受精能力に影響し,精巣ガンおよび乳ガンの発生率を増加させると疑われている物質である。
- 異種ホルモン
- ダイオキシン [こちらも参照のこと]
- グリコール・エーテル
ここでは,それらの作用の分子機構を概説し,生殖システムに生じる影響について述べる。
第1の原因物質: 異種ホルモン xenohormone
異種ホルモンとは ステロイドホルモン 作用またはその抑制作用を示す合成物をいう。それらは
- 農薬 ( エンドスルファン,p,p’DDE,o,p DDT,クロルデンなど )
- 染料および塗料 ( フェノールレッドなど )
- プラスチック材料の可塑剤 ( ビスフェノール A など )
のような広範囲に使用されている化学物質である。あるいはホルモン作用をもつ生体異物の最も良く知られている例の 1 つが,
- ジエチルスチルベストロール( DES )の作用である。
DES の例により,生体異物がホルモン作用を誘発することが初めて証明された。
妊婦に対する DES 処置は,生まれた女性に生殖器のガンならびに奇形の発生率の増加を引き起こした。また,胎児期に DES の影響を受けた男性の中で精子の数や品質が低下することが報告されている。DES は,マウスでも同様の有害効果を引き起こす。
| 【注】DDT=dichloro-diphenyl-trichloro-ethane ; dichlorodiphenyltrichloroethane,ジクロロ・ジフェニル・トリクロロ・エタン,DDT ◆非常に有効な殺虫剤。スイス人科学者 Paul Muller が作った。1940 年代から使われ始めた。しかし人体・自然界に有害で,1972年に米は使用禁止。第 2 次大戦後の日本で米進駐軍が大量に使用。◆【参照】Rachel Louise Carson 沈黙の春。 |
1. 異種ホルモンの作用機序
図1 には正常なステロイドホルモンの作用機序(左)と異種ホルモンの秩序のない作用機序(右)を示した。 異種ホルモンはステロイドホルモンの核受容体,すなわち
- エストロジェン受容体 ( ER )
- アンドロジェン受容体 ( AR )
に作用して,エストロジェンと類似の作用を示したり,テストステロン作用に拮抗したりして,FSH と LH の分泌を変化させて,受胎能力に影響してしまう。
また,テストステロンの合成に関わる重要な酵素反応を抑制する( テストステロン産生を抑制する ),あるいは逆にテストステロンを分解してしまう。
2. 異種ホルモンの影響
( 1 ) エストロジェン受容体を秩序なく活性化してしまう(図1)
- ポリ塩化処理された “農薬”( エンドスルファン,トキサフェン,ディレドリン,o,pDDTなど )は エストロジェン受容体 ( ER ) に直接作用し,
- エストラジオール-17β をその受容体から解離させてしまう。
- エストロジェン受容体 – “農薬” 複合体は,エストラジオールが存在しなくても プロモーター を活性化してしまう( 図 1 )。
エストラジオールと類似の効果をもつ β-ヘキサクロロシクロヘキサン( HCH ) はプロモーター( estrogen responsive element, ERE と呼ばれる )を活性化することはないので,まだ知られていない機序があるものと考えられる。

図1 の正常なステロイドホルモンの作用機序(左)の説明
- 内分泌細胞で分泌されたエストロジェンやテストステロンは”標的細胞” に運ばれる。
- ステロイドホルモンは分子量が小さく,脂溶性分子なので細胞膜を自由に通過できる。
- ステロイドホルモンは細胞質を通過して核に到達する。
- 核内の受容体に結合して生じたステロイドホルモン-受容体複合体は,エストロジェン ( またはテストステロン ) リスポンス・エレメント と呼ばれる特異的な DNA 配列に結合する。
- その結果,特異的な遺伝子が活性化し,mRNA の転写が開始し( 場合によっては,終了し ) ,
- 特異的なタンパク質の合成(または停止)によって,
- ホルモンに対する反応が生じる。
図1 の異種ホルモンの秩序のない作用機序(右)の説明
- 体内に取り込まれた異種ホルモンは血流によって,ステロイドホルモンの”標的細胞” に運ばれる。
- 異種ホルモンは分子量が小さく,細胞膜を自由に通過できる。
- 異種ホルモンは細胞質を通過して核に到達する。
- ステロイドホルモンの代わりに受容体複合体を形成してしまう。
- その結果,本体必要のない特異的な遺伝子が活性化してしまい,その mRNA の転写が開始し( 場合によっては,終了し ) ,
- 無秩序な特異的タンパク質の合成(または停止)によって,
- 正常とは異なる内分泌環境が生じていまう。
( 2 ) アンドロジェン作用または抗アンドロジェン作用を示す(図2)
 o,p DDT のような化合物や,その代謝産物である p,p’ DDE は抗アンドロジェン作用を示す( 図 2 )。
o,p DDT のような化合物や,その代謝産物である p,p’ DDE は抗アンドロジェン作用を示す( 図 2 )。
これらの物質はアンドロジェン受容体 ( AR ) に結合し,その機能を抑止してしまう。性成熟期または成熟後にこれらの化合物の影響を受けると,雄の性徴発現が遅延する。
逆にテストステロンに成り変わって アンドロジェン受容体 ( AR ) に作用してしまう場合があり( ヒドロキシフルタミドなど ),前立腺ガンのようなホルモン依存性ガンの発生への影響が懸念される。
( 3 ) LH と FSH を抑制してしまう(図3)
 LH,FSH およびテストステロンは精子形成を制御する主要なホルモンである。
LH,FSH およびテストステロンは精子形成を制御する主要なホルモンである。
それらの作用は,
に関連して起こる。
- LH – ライディッヒ細胞 – テストステロン分泌軸が抑止されると,精子形成に影響する。
- セルトリ細胞は生殖細胞分化のために必要な支援細胞である。セルトリ細胞の増殖が十分進行しないと,精子形成 に影響する。
( 4 ) ミューラー管の退化が遅延する(図4)
 DES で処理された雄胎子では ミューラー管 が完全に退縮していない( DES が抑制作用を示す)。異種ホルモンが,以下の機序により害作用を及ぼす。
DES で処理された雄胎子では ミューラー管 が完全に退縮していない( DES が抑制作用を示す)。異種ホルモンが,以下の機序により害作用を及ぼす。
- ミューラー管抑制ホルモン を分泌するはずのセルトリ細胞の発育が阻害される。[ ミューラー管抑制ホルモンについてはこちら ]
- ミューラー管抑制ホルモン の分泌自体が抑制される。
- ミューラー管細胞のエストロジェン受容体に直接作用してしまうため,ミューラー管の退化が遅延する。
また,胎児期に作用した DES によって,子宮の奇形も起こる。
第 2 の原因物質:ダイオキシン類
ポリ塩化ジベンゾ-p-ダイオキシンのようなハロゲン化芳香族炭化水素,ビフェニル,ジベンゾフランおよび関連する合成物は持続性をもち,広範囲に及ぶ環境汚染物質の代表である。
ダイオキシン類のうちで最も毒性の強い,2,3,7,8-四塩化ジベンゾ-ダイオキシン( TCDD,ダイオキシン )は,
- ガン化促進
- 免疫障害
- 肝障害
- 皮膚毒性
- 致死性
- 出生異常
- 内分泌攪乱
- 酵素の発現
たとえば,- ミクロソーム内のシトクロム cytochrome P4501A1 ” CYP1A1 “
- アリル・ハイドロカーボン・ヒドロキシラーゼ aryl hydrocarbon hydroxylase ” AHH ” など
等々多種多様な影響をもたらす。
P4501A1アイソザイムは,多環の芳香族炭化水素 ( これらの多くは発ガン物質である ) の代謝活性に関与する。
種々の動物実験の結果から,TCDD は胎子-胎盤障壁を通過し,胎子にまで到達してしまい,発育途上の生殖システムに悪影響を引き起こすことが知られている。
1. ダイオキシンの作用機序
 TCDD の作用は 2 つの転写調節因子,すなわち aryl hydrocarbon receptor ( AhR ) と核への転移因子 Ah receptor nuclear translocator ( ARNT ) の働きで発現する ( 図 5 )。
TCDD の作用は 2 つの転写調節因子,すなわち aryl hydrocarbon receptor ( AhR ) と核への転移因子 Ah receptor nuclear translocator ( ARNT ) の働きで発現する ( 図 5 )。
- TCDD が AhR 受容体に結合すると,
- 酵素 AHH 活性に影響するだけでなく,胸腺退縮や表皮の角質化のような害作用を引き起こす。
- TCDD と結合した AhR は他のタンパク質 ( hsp90 = 90 kDaの熱衝撃蛋白質など ) を解離し,核に移動する。
- DNA に結合する前に,AhR はタンパク質因子 ARNT と結合する。
- AhR-ARNT のヘテロ二量体は,チトクロームP4501A1 ( CYP1A1 ) 遺伝子プロモーター領域にある dioxin responsive element ( DRE ) あるいは xenobiotic response element ( XRE ) と呼ばれる特異的な DNA 配列に結合する。
- これらの相互作用により CYP1A1 遺伝子の転写率が向上して,CYP1A1 の mRNA が蓄積し,タンパク質( 酵素 )が生産される。
| 遺伝子の発現についてはこちら |
2. ダイオキシンの影響:内分泌攪乱物質
( 1 ) LH のフィードバックループを妨げる
TCDD は 顆粒層細胞 の中でいくつかの経路を抑制する。
- ステロイド 合成
- MAP キナーゼ
- 細胞増殖
- プロテインキナーゼ A
- グルコース輸送経路
である。
ラットでは,TCDD により排卵が抑止され,卵巣重量が減少した。上述の異種ホルモンと異なり,TCDD は LH のフィードバックループを妨げるため,結果的に血液中の LH および FSH 濃度を増加させる。
( 2 ) 抗アンドロジェン作用を示す
p,p’-DDE を投与すると,精巣,精巣上体ならびに前立腺の重量が低下する。さらに,TCDD は胎子前立腺におけるステロイド合成酵素( 5α-reductase )を抑制する。
| 精巣 testis; 精巣上体 epididymis; 前立腺 prostate gland |
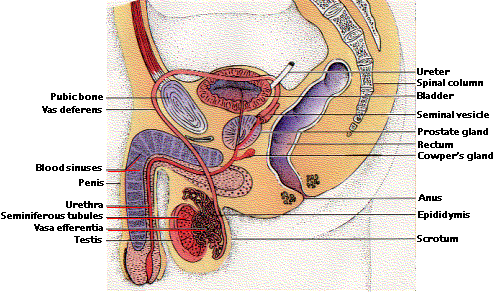{kind=link}
( 3 ) 抗エストロジェン作用を示す
TCDD はマウスならびにラットの乳腺または子宮に対するエストラジオール-17βの作用を抑制する。その他, 精巣上体,前立腺および精巣重量の低下 が観察されている。
TCDD の抗エストロジェン作用は,次の 3 つの機序による。
- エストロジェン受容体の合成を抑制
- エストロジェン受容体のプロモータ領域( ERE )への結合を抑制
- コアクチベータとコリプレッサーとの無差別な相互作用
( 4 ) 抗プロジェステロン作用をもつ
TCDD への長期間暴露によって,アカゲザルでは子宮内膜症が起こり,TCDD によって黄体ホルモン作用が抑制される可能性が示唆されている。これに対し,子宮内膜症に対し,TCDD は影響しないとの報告もあるので,今後の検討が必要である。
第 3 番目の原因物質:グリコール・エーテル

( 1 ) グリコール・エーテルの代謝と作用
グリコール・エーテルは無色の有機溶剤の 1 種で,水および多くの有機溶剤と混和する。このため,ラッカー,塗料,染料,インク,洗浄剤ならびに液体洗剤の製造工程で使用されている。
グリコール・エーテルは,
- エチレン・グリコールファミリー ( E グループ )
- プロピレン・グリコールファミリー ( P グループ )
に分類される。
吸入によるグリコール・エーテルの留保率は非常に高く(すなわち,2-methoxyethanol ( MG ) で70%,butylglycol ( BG ) で57% ),また皮膚からの吸収率も高い。MGは胎児-母胎の障壁を急速に通過してしまう。
グリコール・エーテルは急速に代謝され,血液中または粘膜内で 2 – 3 分以内にヒドロキシ化され,通常それらは体液中に検知されない。しかし,その代謝産物がいろいろな影響を及ぼす。
( 2 ) ヒトと動物のグリコール・エーテル汚染の可能性
動物実験では,影響を受けやすい細胞は 精祖細胞 と同様に,分裂中の 精母細胞 である。E グループの代謝産物,すなわちメソキシ酢酸 ( MAA ) とエソキシ酢酸 ( EAA ) が最も精子形成に毒性を示す。
ヒト尿中のEAAを測定すると,不妊の男性で 3.8% と高濃度の EAA が検出されている( 正常男性の場合 1.2% )。
プラスチックとインクの生産に使用されるフタル酸エステル ( DEHP ) などの化合物は,実験動物の精巣萎縮を引き起こすが,ヒトではこの影響に関する情報はない。米国での疫学的調査 ( 1980 – 1989 ) では,エチレン・グリコール・エーテルに曝された女性が,自然流産および受胎率低下の割合が高いことが示された。
これらの結果を受け,卵母細胞に対するエチレン・グリコール・エーテルの影響がラットで検討されたところ,
- 排卵数の減少
- 体外受精率の低下
が示された。
中~高濃度のプロピレン・グリコール・モノメチル・エーテルに暴露させた雌ラットについて行われた研究では,
- 受胎率の低下
- 発情周期 の延長
- 卵巣重量の低下
- 卵巣萎縮
が示されている。
動物実験の結果は,ヒトで報告されている観察と一致している。したがって,グリコール・エーテルはヒトの生殖に重大な脅威をもたらすものと思われる。
( 3 ) グリコール・エーテルは核( DNA )に影響する
グリコール・エーテル由来のアルデヒドおよび酸の代謝産物が 細胞周期 を混乱させ,染色体異常などを起こす。低濃度の methoxyaldehyde ( 0.07 mM ) で処理された細胞では,染色体が分散したり紡錘極が消失した。
これらの結果は,グリコール・エーテルが生殖細胞の DNA 傷害を引き起こす可能性を示唆する。したがって,グリコール・エーテルに曝された人における自然流産,奇形および精巣ガンとの関連が疑われる。
( 4 ) 遺伝子調節に対するグリコール・エーテルの影響
遺伝子調節に対するグリコール・エーテルの影響を検討した最近の研究では,遺伝子の発現が変化していることが見出されている。
| 遺伝子の発現についてはこちら |
疫学的な分析データ,動物上の生体内と生体外の実験データならびに分子生物学的データが,グリコール・エーテルの生殖毒性を指し示している。
この化合物は異なる標的細胞に作用して,
- 細胞 アポトーシス
- ガン化
- 不妊
- 奇形
なぜこれらの化合物がホルモン以上に危険なのか?
- 生体内由来のホルモンが分泌されていないか,あるいはまた不活性な発育時期に,これらの化合物による汚染が起こることがある。
この場合は,必要のない時期または場所で,ホルモン受容体の無秩序な活性化が起こってしまうので,発生上の奇形が生じる可能性がある。 - 生体内由来のステロイドホルモンは血清蛋白質に結合して輸送され,かつホルモン濃度が調整される。
しかし,異種ホルモンは血清蛋白質とほとんど結合せず,遊離した状態のためそれらの濃度は常に高く,濃度調節のないまま作用が引き起こされてしまう。 - シトクロムP450 (水酸化酵素ファミリーの総称)が関与して,ステロイドホルモンなどの脂溶性の分子を水酸化して,排出されやすい水溶性の物質に変える。
しかし,生体異物ははるかに高い安定性をもつため,長期間これらに暴露されることになり,累積的な有害作用が生じてしまう。
| 最初に戻る |
| メニューのページへ戻る |
February 05, 2020